Что это — патогенный гриб?
Грибы представляют собой отдельное царство животного мира. Они имеют множество форм: съедобные, ядовитые, плесень, дрожжи и многие другие. Современной науке известно больше пятисот видов грибов. Эти создания встречаются повсюду на нашей планете, даже внутри человека. Некоторые из них хорошо уживаются с людьми и составляют условно-патогенную микрофлору. Патогенный гриб обязательно вызывает заболевание. Он потворствует своей природе и стремится отвоевать себе место под солнцем, а также ресурсы для дальнейшего роста и развития. К сожалению, это вредит здоровью человека.
Определение
Патогенные грибы – это возбудители глубоких и поверхностных микозов у человека и животных. Эти создания относятся в основном к классу дерматофитов, то есть питающихся кожей. Реже среди них встречаются низшие грибы и актиномицеты.
У них существует определенная тропность к тканям животных. Это значит, что дерматофиты предпочитают эпидермис с волосистую часть кожи, дрожжи – лимфатическую систему, кандиды – паренхиматозные органы, аспергиллы живут в дыхательной системе, а актиномицеты обожают селиться в костях.
Зная эти особенности врач может дифференцировать заболевания и назначать специфическое лечение.
Классификация патогенных грибов
В царстве грибов патогенные грибы делятся на два отдела: слизевики и настоящие грибы. Последний разбирается на семь классов, названия которых отражают присущие им стадии развития:
— цитридомицеты;
— гипоцитридомицеты;
— оомицеты;
— зигомицеты;
— аскомицеты;
— базидомицеты;
— дейтеромицеты.
Первые четыре представителя формируют группу низших грибов, остальные относятся к высшим, а последний класс — к несовершенным грибам. Большинство патогенных грибов, вызывающих заболевание у людей, относятся к дейтеромицетам.
Свойства патогенных грибов
Человек обычно сразу не замечает, что в его организм попали патогенные грибы. Споры (семена грибов) удлиняются и принимают вид трубки, которая продолжает расти и истончаться, чтобы со временем превратиться в гифу и стать основой грибницы. Уже на этом этапе заметно отличие. Гифа высших грибов имеет перегородки, а низших – нет. Гифы от разных спор растут, сплетаются между собой и в конечном итоге на субстрате вырастает мицелий.
Уже на этом этапе заметно отличие. Гифа высших грибов имеет перегородки, а низших – нет. Гифы от разных спор растут, сплетаются между собой и в конечном итоге на субстрате вырастает мицелий.
Для диагностики и производства лекарств патогенные виды грибов выращивают на питательных средах, таких как Сабуро, Чапека-Докса, на сусле и сусло-агаре. Обязательным условием является рН ниже семи.
Клетки грибов покрыты стенкой из углеводов, но веществом, по которому можно определить видовую принадлежность, остается хитин. Он не взаимодействует с пенициллинами и лизоцимом, поэтому обладает большей вирулентностью для человеческого организма.
Патогенный гриб устойчив к физическим и химическим дезинфектантам. Лечение от них способно нанести непоправимый вред органам и системам человека, так как требуется высокая концентрация препаратов в жидкостях организма. Наиболее чувствительны к терапии микроспоры, а наименее – кандиды. Подбор препаратов усложняется тем, что у одного вида грибов возможны различные комбинации антигенов, а токсины, ферменты и другие факторы патогенности до сих пор остаются неизвестными.
Особенности инфекции у человека
Грибы, патогенные для человека, способны вызвать заболевания, которые можно разделить по локализации на четыре группы:
- Глубокие микозы – это поражение паренхиматозные органов, сепсис, диссеминация спор из очага заболевания в соседние ткани.
- Подкожные микозы, они же субкутанные. Грибы заселяют эпидермис, дерму, подкожно-жировую клетчатку, фасции и даже кости.
- Эпидермомикозы или дерматомикозы возникают на производных верхнего слоя кожи: волосах и ногтях.
- Поверхностные микозы (кератомикозы). Патогенные грибы на коже поражают только роговой слой и волосы.
Отдельной группой выделяют заболевания, возбудителями которых являются условно-патогенные грибы. Это оппортунистические заболевания, которые появляются, когда иммунная защита организма ослабляется, например ВИЧ, гепатит В или С, онкологическое заболевание.
Чаще всего возбудители микозов находятся в почве или пыли, поэтому важно работать в респираторах, мыть овощи и зелень, проводить влажную уборку в помещениях. Глубокие микозы появляются после вдыхания возбудителя, а для развития кожных заболеваний необходимо, чтобы споры попали на раневую поверхность.
Иммунитет
Патогенный гриб, попадая в организм, вызывает каскад реакций иммунной системы, необходимых для того, чтобы идентифицировать антиген и выработать против него специфическую защиту.
Как правило, все грибы — сильные иммуногены, поэтому у людей часто имеется аллергия на них. Реакция развивается по типу гиперчувствительности замедленного типа или цитотоксическому типу. Помимо этого Т-хелперы стимулируют тканевые макрофаги для элиминации спор. Гуморальные реакции проявляются в виде высокого титра антител, по которому можно определить стадию развития инфекции, а также в виде активации системы комплемента по классическому и альтернативному пути.
Диагностика микозов
Самый простой способ выявить патогенный гриб – микроскопия. У пациентов берут кровь, слизь и кожу с пораженных участков, наносят на предметные стекла, окрашивают или обрабатывают кислотами, а затем помещают в световой или электронный микроскоп. Такая процедура позволяет рассмотреть морфологические особенности возбудителя и определить его вид.
Иногда в лаборатории высеивают грибы на селективные среды и наблюдают на их ростом и ферментированием различных веществ. Это помогает с биохимической точки зрения определить возбудителя.
В ответ на внедрение патогенных грибов в крови человека появляются антитела, факт наличия которых можно определить серологическими методами исследования. Однако результат такой процедуры может быть неточным, так как разные виды грибов содержат перекрестно реагирующие антигены.
В эпидемиологических исследованиях, для того чтобы выявить ту часть популяции, которая уже переболела грибковыми инфекциями, использовали кожные пробы. Это позволяло узнать, сталкивался ли ранее организм с этим видом антигенов или нет. Для диагностики этот метод использовать нельзя, так как он имеет низкую специфичность.
Это позволяло узнать, сталкивался ли ранее организм с этим видом антигенов или нет. Для диагностики этот метод использовать нельзя, так как он имеет низкую специфичность.
Род кандида
На сегодняшний день выделено 186 видов рода кандида, но только некоторые из них могут вызывать заболевания у человека. Например, C. albicans, C. pseudotropicalis, C. tropicalis, C. krusei, C. parapsilosis, C. Quillermondii и другие.
Это условно-патогенные грибы, которые постоянно находится в кишечнике человека. Хорошо растут на средах, богатых углеводами. Колонии состоят их мелких овальных клеток, переплетенных с нитями мицелия. Очень быстро размножаются в крови при нормальной температуре в 37 градусов, уже за три часа из нескольких спор образуются тысячи новых гиф. Прорастание клеток в ткани сопровождается сильной местной иммунной реакцией с образованием гноя.
У здорового человека и животных в ротовой полости в 50 процентах случаев высеваются грибы рода кандида, в фекалиях – практически всегда, на коже и слизистой половых путей – до 10 процентов. Разовьется ли заболевание, во многом зависит от состояния иммунной и эндокринной систем. Спровоцировать кандидоз может лекарственная терапия иммуносупрессорами, глюкокортикостероидами, цитостатиками, радиационная болезнь, длительное лечение антибиотиками, онкологические заболевания и прием оральных контрацептивов.
Патогенные грибы вызывают заболевания на фоне сахарного диабета, нарушения функции желез внутренней секреции и других. С недавнего времени значительно возросло количество ятрогенных кандидозов после оперативных и диагностических вмешательств. Кроме того, поражение кожи и слизистых грибом рода кандида является одним из маркеров СПИДа.
Пневмоцистная пневмония
Pneumocystis carinii – гриб, который преимущественно поражает ткани дыхательной системы. Для того чтобы посмотреть на его культуральные свойства, недостаточно обычных питательных сред, необходимо использовать куриные эмбрионы или перевиваемые культуры клеток.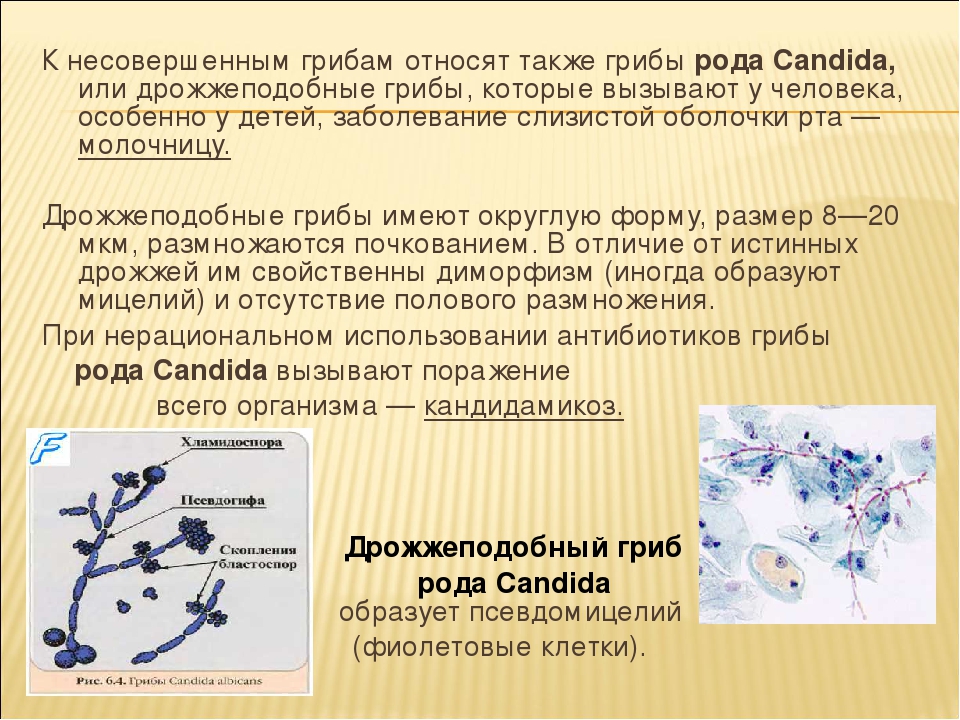
Цисты представляют собой круглые клетки, внутри которых видны базофильные тельца. В колонии вокруг зрелых цист всегда располагаются молодые и промежуточные формы. Наличие внутриклеточных телец позволяет ученым отнести пневмоцисты к классу актиномицетов.
Эти грибы вызывают пневмонию, но в некоторых случаях возможно поражение и других внутренних органов: почек, селезенки, лимфатической системы, сетчатки глаза, сердца, печени, поджелудочной и даже головного мозга. Инфекция, как правило, возникает у детей на фоне сниженного иммунитета.
Аспергиллез
Этот гриб образует гладкие зеленые колонии, которые хорошо растут при температуре тела человека, но плохо переносят жару. Часто встречаются в пищевых продуктах, древесине. Вызывают острую инфекцию после попадания большого количества спор в организм человека вместе с пищей, например хлебом. Часто заболевание развивается вторично, на фоне патологий крови, саркомы, туберкулеза, терапии кортикостероидами, иммуносупрессорами. Не передается от человека к человеку.
Чаще всего поражает дыхательную систему, иногда вызывает болезни кожи, такие как экзема. Вокруг мицелия ткани некротизируются, в очаге поражения появляются гранулемы. Характерным признаком является возникновение полостей на пораженных участках, которые содержат грибковые шары. В литературе описаны случаи генерализованной инфекции с поражением центральной нервной системы.
12.Грибы, классификация, патогенные и условно-патогенные виды, методы выявления.
Грибы
относятся к царству Fungi (Mycetes, Mycota). Это
многоклеточные или одноклеточные
нефотосинтезирующие (бесхлорофильные)
эукариотические микроорганизмы с
клеточной стенкой.
Грибы
имеют ядро с ядерной оболочкой, цитоплазму
с органеллами, цитоплазматическую
мембрану и многослойную, ригидную
клеточную стенку, состоящую из нескольких
типов полисахаридов, а также белка,
липидов. Цитоплазматическая мембрана
Цитоплазматическая мембрана
содержит гликопротеины, фосфолипиды и
эргостеролы. Грибы являются
грамположительными микробами, вегетативные
клетки — некислотоустойчивые. Тело
гриба называется талломом.
Классификация
грибов.
Различают
два основных типа грибов: гифальный
и дрожжевой.
Гифальные
(плесневые)
грибы образуют ветвящиеся тонкие нити
(гифы), сплетающиеся в или мицелий. Гифы
низших грибов не имеют перегородок.
Они представлены многоядерными клетками.
Гифы высших грибов
разделены
перегородками с отверстиями.(совершенные
грибы)
Дрожжевые
грибы
имеют
вид отдельных овальных клеток.
Одноклеточные грибы, которые по типу
полового размножения распределены
среди высших грибов — аскомицет и
базидиомицет. При бесполом размножении
дрожжи образуют почки или делятся, что
приводит к одноклеточному росту.
Среди
грибов, имеющих медицинское значение,
выделяют 3 типа: зигомицеты (Zygomycota),
acкомицеты (Ascomycota) и базидиомицеты
(Basidiomycota).
Зигомицеты:
низшие грибы, вызывают зигомикоз легких,
головного мозга.
Аскомицеты:
высшие совершенные грибы, к ним относится
возбудитель эрготизма, паразитирующий
на злаках. Микозы, дерматомикозы,
пневмония, кератоз, аспергиллез.
Базидиомицеты:
шляпочные грибы. Болезни: отравление
ядовитыми грибами, криптококкоз.
Дейтеромицеты:
несовершенные грибы, род Candida, поражающие
слизистые оболочки и внутренние органы.
Заболевания:
1.
Поверхностные микозы (кератомикозы) —
поражения поверхностных слоев кожи и
волос:Malassezia furfur, Cladosporium.
2.Эпидермофитии
(эпидермомикозы) — поражения эпидермиса,
кожи и волос:Epidermophyton floccosum,Microsporum canis.
3.Подкожные,
вовлекающие в процесс дерму, подкожные
ткани, мышцы: Sportrichum schenckii.
4.Системные,
или глубокие, микозы, при которых
поражаются внутренние органы и ткани:
Coccidioides immitis, Histoplasma capsulatum/
5.
Оппортунистические микозы: Candida,
Aspergillus, Mucor. (усл-пат)
6.
Аллергии, вызванные грибами (пневмоаллергии
и дермоаллергии).
7.
Микотоксикозы
—
пищевые интоксикации, вызванные токсинами
грибов.
Лабораторная
диагностика:
микроскопическое, микологическое,
аллергическое, серологическое
исследования.
Материал
для исследования: гной, мокрота, волосы,
кожа, кровь.
Микроскопическое:
микроскопия нативных препаратов:
изучение строения гриба, расположение
спор, культуральная принадлежность.
Для
окраски – методы Грамма, Романовского,
Циля-Нильсена.
Культуральное
(микологическое) исследование: выделение
чистой культуры, плотные, жидкие пит.
среды.
Серологическое
реакции: для диагностики грибковых
заболеваний, с грибковыми АГ. (РА,
РСК,РНГА,РИФ).
Аллергические
пробы – в/к введение аллергенов.
Лечение:
поверхностных микозов: удаление
пораженных участков с помощью
кератолитических средств. Препараты,
содержащие дисульфид селена, тиосульфат.
Эпидермофитии: противогрибковая терапия,
гризеофульвин, антимикотики. Кетоконазол.
Амфотерицин В. При кандидозе: нистатин,
леворин.
5. Патогенные грибы
Дрожжеподобные
грибы Кандида –
это широко распространённые в природе
микроорганизмы, которые не способны
вызывать спиртовое брожение, однако,
они способствуют порче пищевых продуктов,
а некоторые их агрессивные формы вызывают
патологические процессы в организме
человека и животных.
Строение и размножение гриба кандида
Грибы
рода Candida имеют
вытянутую цилиндрическую форму и
вопределённых условиях способны
образовывать примитивный мицелий.
Наиболее
распространёнными представителями
данного рода являются Candida spp. (Candida
ablicans, Сandida tropicalis, Сandida pseudotrocalis и Сandida
krusei).
Грибок
кандида не
образуют истинных спор (эндоспор),
которые характерны практически для
всех сумчатых грибов, к которым относится
род Кандида. Однако, при возникновении
неблагоприятных условий, эти микроорганизмы
способны формировать хламидоспоры
(несколько видоизменённая клетка,
которая покрыта плотной оболочкой). Как
правило, хламидоспоры являются одним
из важнейших факторов заражения.
Псевдомицелий
кандиды состоит из нескольких удлинённых
клеток, которые соединены в длинные
цепочки. Необходимо отметить, что каждая
клетка при этом имеет свою отдельную
полноценную клеточную мембрану, как и
истинный мицелий.
Размножение
происходит в виде почкования. При этом
от материнской клетки отделяется
бластопора (новый клеточный зачаток),
который со временем превращается в
полноценную взрослую клетку.
Среда для развития кандиды
Наиболее
благоприятным условием для существования
гриба кандиды является слабощелочная
или нейтральная среда. При этом необходимо
отметить, что даже незначительное
смещение среды в кислую сторону
значительно подавляет развитие и
размножение возбудителя. В ходе своей
жизнедеятельности гриб
кандида способна
сдвигать кислотный баланс в сторону
слабощелочной реакции, в связи с чем,
вызванная им патология носит лавинообразный
характер.
В процессе
роста патогенных грибов на питательных
средах в них постоянно происходят
различные химические процессы: прививки,
сообщения, образования органических
кислот, стеринов, полисахаридов,
витаминов, пигментов и другие. Вследствие
этого меняются не только культуральные
и морфологические условия, но и условия
спорообразования и характер образования
мицелия.
Для
полноценного питания и роста все виды
грибов нуждаются в целом ряде химических
элементов, в частности K, Na, Cu, P, C, H, O, N,
Mg, Mn, Fe, Zn и т. п. Потребность в углероде
п. Потребность в углероде
грибы удовлетворяют за счет органических
соединений. Кислород грибы берут, как
правило, из воздуха, а также, вероятно,
из питательной среды. Азот получают из
органических веществ: белков, пептонов,
аминокислот. Жировой обмен у патогенных
грибов изучен очень мало. Известно, что
для роста некоторых групп и видов грибов
нужны жирные кислоты. Плесневые грибы
обладаютособой способностью расщеплять
жиры. Рядом исследователей была обнаружена
липаза в плесневых грибах рода Penicillium,
Aspergillus flavus.
Кроме
того, в результате исследования жирового
обмена в человеческой коже было
обнаружено, что в расщеплении жиров
зачастую участвуют грибы рода Нyphomycetes,
а в расщеплении лецитина — грибы, которые
размножаются почками. Важное значение
для жизнедеятельности патогенных грибов
имеют отдельные витамины. По способу
добывания этих жизненно важных веществ
грибы делятся на две большие группы. В
частности, нитчатые грибы размножаются
почками и вполне самостоятельно
обеспечивают себя витаминами, синтезируя
их из питательной среды.
Другие
группы грибов не могут добывать себе
достаточное количество нужных витаминов.
Поэтому их принято называть грибами с
«недостатком витаминов». Большинство
грибов постоянно производят энзимы, а
некоторые виды проявляют эту способность
только при определенных условиях
существования. Энзимы, вырабатываемые
в процессе такого приспособления,
называются адаптивными энзимами.
Дерматомицеты способны расщеплять
растительные и животные жиры, а из
продуктов распада ассимилировать не
только глицерин, но и различные жирные
кислоты. Большинство дерматомицетов
также расщепляют холестерин.
Одним
из ведущих свойств дрожжеподобных
грибов является способность расщеплять
виноградный сахар и другие виды сахара
на этиловый спирт и углекислый газ с
помощью производимых ими энзимов.
Многочисленные виды грибов производят
энзим амилаза, который разлагает крахмал.
Из культур некоторых грибов, в частности
родов Aspergillus и Penicillium, можно выделить
пектиназу, проявляющую катализирующие
действие при расщеплении пектина.
Нитчатые и грибы, которые размножаются
почками, поражают коллаген и эластин.
Кроме того, нитчатые грибы способны
разлагать кератин.
★ Условно патогенные грибы | Информация
Пользователи также искали:
классификация патогенных грибов,
патогенные грибы микозы,
патогенные грибы на ногте,
патогенные грибы споры обнаружены,
патогенные грибы в мазке,
споры патогенного гриба на коже,
виды грибов патогенных для человека,
споры обнаружены,
классификация патогенных грибов,
на ногте,
микозы,
споры патогенного гриба на коже,
виды грибов патогенных для человека,
в мазке,
патогенные грибы споры обнаружены,
патогенные грибы на ногте,
патогенные грибы микозы,
патогенные грибы в мазке,
разные,
условно,
патогенные грибы,
грибы,
патогенные,
патогенными грибами,
патогенный гриб,
патогенных грибов,
патогенных,
условно патогенных грибов,
патогенную,
патогенной,
патогенная,
гриб,
условно патогенные,
грибами,
грибов,
патогенным,
патогенными,
Патогенные грибы
Содержание:
- Морфология
грибов.
- Классификация
возбудителей микозов. - Кокцидиоидоз
. - Гистоплазмоз.
- Актиномикоз.
- Нокардиоз,
мадуромикоз.

7. Литература.
Грибы являются
эукариотами и относятся к низшим растениям.
Они отличаются как своим более сложным
строением, так и более совершенными способами
размножения. Среди большого разнообразия
грибов, приспособившихся к жизни в почве
и воде, выделяется группа паразитических,
вызывающих заболевания человека, животных,
птиц и поражающих растения. Заболевания
эти получили название микозов (от mycetes
— грибы).
Клетки грибов
— гифы — покрыты оболочкой, имеют
ядро с ядрышком, цитоплазму и включения.
Размножение грибов
может происходить двумя
половым и бесполым. К совершенным
относятся те, для которых характерны
оба пути размножения; несовершенными
называются грибы, у которых нет или еще
не описан половой путь размножения. Бесполое
размножение осуществляется у низших
грибов с помощью эндогенных спор, созревающих
внутри круглой структуры — спорангия,
и экзогенных — конидий, формирующихся
на кончиках плодоносящих гиф. Возможен
и третий путь размножения — вегетативный
— при почковании клеток или оторвавшимися
участками мицелия.
Согласно ботанической
классификации, грибы подразделяются
на две категории: низшие, к которым относятся
классы архимицеты (Archimycetes) и фикомицеты
(Phycomycetes), и высшие, включающие также два
класса — аскомицеты (Ascomycetes) и базидиомицеты
(Basidiomycetes). В пятый класс — дейтеромицеты(Deuteromycetes),
или несовершенные грибы (Fungi imperfecti), объединены
грибы, у которых не описан половой путь
размножения. Самостоятельную группу
Самостоятельную группу
составляют дрожжи и дрожжеподобные грибы.
Возбудители микозов
относятся к различным классам.
1. Архимицеты.
Среди этого класса грибов патогенных
видов для человека нет. По своему строению
это наиболее просто организованные грибы;
большинство из них являются внутриклеточными
паразитами водорослей. Клетки этого класса
грибов редко образуют мицелий и являются
одноядерными небольшими гифами.
2. Фикомицеты.
Объединяют большую группу грибов, имеющих
несептированиый хорошо развитый мицелий
и различающихся по способам полового
размножения. Среди фикомицетов выделяют
хитридиомицеты (Chitridiomycetes) или водные
грибы, как ведущие сапрофитический образ
жизни, так и вызывающие поражения водорослей,
водных растений. Группа оомицетов (Oomycetes)
объединяет облигатных паразитов высших
растений (наземные формы) и водяные плесени
(свободноживущие в воде подвижные формы).
3 и гоми цеты (Zygomycetes) также являются наземной
группой грибов класса фикомицетов. Среди
них хорошо изучена плесень Mucor mucedo (рис.
11), широко распространенная в почве и
воздухе; некоторые патогенные виды ее
могут вызывать заболевание у человека
— мукороз. При бесполом размножении плесени
на плодоносящей гифсспорангиеноспе образуется
мешокспорангий круглой формы, внутри
которого созревают споры. При созревании
спорангий лопается, эндоспоры высыпаются;
каждая спора дает начало мицелию.
3. Аскомицеты.
Объединяют группу высших грибов, имеющих
септированный мицелий и отличающихся
особенностью полового размножения. Свое
название аскомицеты получили от основного
органа снороношения — аска (сумка), являющегося
зиготой. Аск содержит 4 или 8 гаплоидных
половых спор. В результате разрыва аска
споры освобождаются и, прорастая, дают
начало новому мицелию. К аскомицетам
относятся грибы родов Aspergillus (рис. 12), Penicillium
(рис. 13), отличающиеся особенностями формирования
плодоносящих гиф. У Aspergillus (леечная плесень)
на концах плодоносящих гиф — конидиеносцах
имеются утолщения — стеригмы, на которых
образуются цепочки экзоснор — конидии.
Стеригма с отходящими от нее конидиями
напоминает струи жидкости, льющиеся из
лейки. Некоторые виды аспергилл могут
вызывать поражения кожи и легких у человека.
Плодоносящая гифа у грибов рода Penicillium
(кистсвик) напоминает кисточку, так как
на конидиеноспс образуются утолщения,
разветвляющиеся на более мелкие структуры
— стеригмы, на которых находятся терминальные
цепочки сферических конидий. Из некоторых
видов зеленой плесени (Penicillium notatum, Penicillium
chrysogenum) был получен первый антибиотик
— пенициллин. Среди пепициллиумов есть
патогенные виды, вызывающие заболевание
у человека, — пенициллиоз.
4. Базидиомицеты.
Шляпочные грибы, съедобные и ядовитые,
а также некоторые паразиты деревьев и
растений. Бесполое размножение происходит
посредством образования конидий; при
половом зигота превращается в булавовидную
клетку — базидию. Ядро, разделив , шесся
внутри базидии на четыре новых, вместе
с частью цитоплазмы отстреливается от
базидии.
5. Несовершенные
грибы. Объединяют более 25 000 видов высших
грибов, у которых отсутствует или еще
не определен половой путь размножения.
К несовершенным грибам относятся многие
патогенные виды, такие, как возбудители
трихофитии, микроспории, парши, эпидермофитии.
Классификация несовершенных грибов основывается
на формах спороношсния и имеет практическое
значение.
6. Дрожжевые
грибы (дрожжи). Одноклеточные организмы,
не образующие мицелий. Клетки имеют овальную
форму, размер 2—20 мкм. Обычно размножаются
почкованием, некоторые виды — бинарным
делением. Большинство видов дрожжей —
сапрофиты; их способность вызывать брожение
широко используется в пищевой и многих
других видах промышленности (хлебопечение,
пивоварение и т. д.). Заболевания, вызываемые
некоторыми видами дрожжей, получили название
дрожжевых микозов.
Дрожжеподобные
грибы. Отличаются от истинных дрожжей
способностью образовывать псевдомицелий:
почкующиеся дочерние клетки не отходят
от материнской, а вытягиваются в нити
(рис. 14). Дрожжеподобные грибы рода Candida
14). Дрожжеподобные грибы рода Candida
относятся к несовершенным грибам. Они
имеют овальную или круглую форму, диаметр
2—5 мкм, делятся почкованием, образуют
псевдомицелий, на концах которого могут
появляться споры, покрытые двойной оболочкой
(хламщоспоры). С. albicans, С. tropicalis и др. вызывают
у человека каидидоз — поражение кожи,
слизистых оболочек и внутренних органов.
Возможно экзогенное заражение при контакте
с больным человеком, но чаще кандидоз
возникает как эндогенная инфекция. Последнее
обстоятельство объясняется тем, что С.
albicans является представителем нормальной
микрофлоры человека и заболевание кандидозом
возникает при нарушении обычных соотношений
в микробных ассоциациях: например, при
длительной и неправильной антибиотикотерапии,
при хронических заболеваниях и т.п.
КРАТКАЯ
ХАРАКТЕРИСТИКА НЕКОТОРЫХ
МИКОЗОВ
Кокцидиоидоз
Coccidioides
immitis — почвенный сапрофит , обитающий
в засушливых областях США,
Мексики, Центральной и Южной
Америки. В пределах США большинство случаев
кокцидиоидоза приходится на Калифорнию,
Аризону и западную часть Техаса. Изредка
встречаются привозные случаи — когда
возбудитель завозится из эндемического
района, например вместе с кипами хлопка.
Заражение
людей и животных происходит воздушно-пылевым
путем — при вдыхании разносимых ветром
артроспор. У 60% больных первичная легочная
инфекция протекает бессимптомно. Клинически
выраженные случаи — острый легочный кокцидиоидоз
— проявляются как легким гриппоподобным
синдромом , так и тяжелой пневмонией .
Легкие
формы острого легочного
привлекают внимание врачей лишь благодаря
вспышкам или появлению аллергических
реакций ( узловатая эритема , полиморфная
экссудативная эритема , артралгия
, полиартрит , конъюнктивит , эписклерит
). Вспышки обычно возникают через 10-14 сут
Вспышки обычно возникают через 10-14 сут
после контакта с зараженной пылью — во
время раскопок, сбора минералов для коллекций,
войсковых учений, строительных работ
и т. п. Пыльные бури разносят споры на
большие расстояния, что может приводить
к вспышкам инфекции и в тех районах, где
Coccidioides immitis не обитает. Острый легочный
кокцидиоидоз обычно проходит самостоятельно.
Очаг в легких либо заживает бесследно,
либо оставляет после себя монетовидный
рубец или, реже, тонкостенную полость,
что знаменует собой переход болезни в
хроническую стадию. В этом случае кокцидиоидоз
протекает как хроническая пневмония
или как фиброзно-кавернозный туберкулез
.
Иногда
единственным проявлением острого
легочного кокцидиоидоза служит
плевральный выпот. Лечения он обычно
не требует.
Редкое,
но крайне опасное осложнение — гематогенная
диссеминация возбудителя. Если инфекция
выходит за пределы легких и прикорневых
лимфоузлов, говорят о диссеминированном
кокцидиоидозе. Он особенно часто встречается
среди негров, филиппинцев, индейцев, мексиканцев,
а также среди лиц, страдающих иммунодефицитами
(в частности, ВИЧ-инфицированных ), и беременных
.
Coccidioides
immitis вызывает гранулематозное
нередко с казеозным некрозом.
Очаги в легких и прикорневых
лимфоузлах иногда обызвествляются. Основную
защитную роль играет клеточный иммунитет.
У заболевших образуются также антитела
классов IgG и IgM, однако защитными свойствами
они, по-видимому, не обладают. Титр IgG-антител
примерно соответствует количеству возбудителя;
высокий титр — плохой прогностический
признак.
Аллергические
реакции замедленного типа на антигены
Coccidioides immitis присущи легким формам болезни,
в частности острому легочному
кокцидиоидозу. У половины больных
диссеминированным
кожные пробы с антигенами Coccidioides immitis
отрицательны, что предвещает неблагоприятный
исход.
Клиническая
картина:
Острый
легочный кокцидиоидоз проявляется
лихорадкой , кашлем , болью в груди
, недомоганием , иногда — аллергическими
реакциями. При рентгенографии грудной
клетки выявляют затемнения , увеличение
прикорневых лимфоузлов , плевральный
выпот . Возможна легкая эозинофилия .
Через 1-2 нед состояние начинает улучшаться
и, как правило, наступает выздоровление.
Образование полостей в легких примерно
в половине случаев клинически не проявляется,
у остальных больных отмечаются кашель
и кровохарканье .
Хронический
легочный кокцидиоидоз сопровождается
продуктивным кашлем , лихорадкой и
похуданием . Первые признаки диссеминированного
кокцидиоидоза, как правило, обнаруживаются
еще во время первичной легочной инфекции.
Изредка диссеминация возникает спустя
годы после заражения, обычно — при сопутствующем
иммунодефиците ( лимфогранулематоз ,
лимфомы , трансплантация почки , ВИЧ-инфекция
). Признаки, на основании которых следует
заподозрить диссеминацию у больного
острым легочным гистоплазмозом , включают
упорную лихорадку и недомогание , увеличение
прикорневых лимфоузлов , увеличение паратрахеальных
лимфоузлов , повышение СОЭ и высокий титр
комплементсвязывающих антител. Со временем
очаги инфекции появляются в костях, коже,
подкожной клетчатке, оболочках мозга,
суставах и других органах. Единственным
проявлением диссеминированного кокцидиоидоза
может быть хронический менингит . При
этом в СМЖ появляются комплементсвя-зывающие
антитела, а результаты посева и микроскопии
СМЖ чаще всего отрицательны. Поражение
кожи представлено безболезненными папулами
или узлами ; поражение костей и мягких
тканей — абсцессами , которые могут прорываться
наружу с образованием свища . Без лечения
болезнь прогрессирует и через несколько
месяцев или лет приводит к смерти.
Диагностика:
При подозрении
на кокцидиоидоз проводят посев и
микроскопию нативных препаратов мокроты,
мочи и гнойного отделяемого из свищей.
Следует обязательно проинформировать
сотрудников лаборатории о своих подозрениях,
так как работа с культурой Coccidioides immitis
требует крайней осторожности. Патогномоничный
признак кокцидиоидоза — обнаружение зрелых
сферул в биопсийном материале; мелкие
сферулы нужно дифференцировать с дрожжевыми
клетками Blastomyces dermatitidis и Cryptococcus neoformans
. В диагностике кокцидиоидоза широко
используются также серологические методы.
При массовых обследованиях применяются
латекс- агглютинация и иммунодиффузия.
У серопозитивных больных для подтверждения
диагноза и определения титра антител
используется реакция связывания комплемента.
Ее результаты, однако, зависят от тяжести
болезни. Положительные результаты встречаются
практически у всех больных с диссеминированным
кокцидиоидозом, в то время как при остром
легочном кокцидиоидозе и одиночных полостях
в легких они редки. В диагностике первичной
легочной инфекции помогает сероконверсия,
однако она может произойти лишь через
8 нед после начала болезни. Обнаружение
комплементсвязывающИх антител в нативной
СМЖ подтверждает диагноз менингита. Правда,
изредка их находят и при расположении
очага вблизи от оболочек мозга. Кожные
пробы с кокцидиоидином и сферулином (двумя
имеющимися в продаже антигенами Coccidioides
immitis ) при первичной легочной инфекции
становятся положительными через 3-21 сут
после начала болезни (положительным результатом
считается появление в месте введения
антигена через 24-48 ч папулы диаметром
не менее 5 мм). Кожные пробы используют
в основном при эпидемиологических исследованиях
(например, во время вспышек или для определения
границ эндемических районов). Диагностическая
ценность этих проб невелика из-за того,
что положительные результаты могут быть
следствием давно перенесенной болезни,
а отрицательные — нередки при диссеминированном
и хроническом легочном кокцидиоидозе.
Лечение:
Острый
легочный кокцидиоидоз обычно разрешается
самостоятельно. При тяжелом или
При тяжелом или
затяжном течении болезни иногда на несколько
недель назначают амфотерицин В или итраконазол
, чтобы предотвратить развитие диссеминированного
или хронического легочного кокцидиоидоза.
При диссеминированном кокцидиоидозе
с тяжелым, быстро прогрессирующим течением
лечение начинают с амфотерицина В в дозе
0,5-0,7 мг/кг/сут в/в, а через 2-3 мес, если состояние
улучшается, переходят на кетоконазол
(400 мг/сут внутрь), итраконазол (200 мг 2 раза
в сутки внутрь) или флуконазол (400-600 мг/сут
внутрь). В более легких случаях можно
сразу начинать с этих препаратов. Поскольку
заболевание часто рецидивирует, требуется
многолетний или пожизненный прием противогрибковых
средств . При менингите обычно назначают
флуконазол , 400-800 мг/сут внутрь или в/в,
однако может потребоваться интратекальное
введение амфотерицина В . Нелеченный
менингит нередко осложняется гидроцефалией
. При поражении костей и мягких тканей
показаны хирургические методы лечения
(дренирование абсцессов, удаление секвестра
и иссечение пораженной костной ткани).
Анализ ногтей и кожи на грибок или патогенные грибы
Дерматомикозы — это грибковые заболевания кожи и ее придатков, к которым относят волосы, ногти. Возбудители дерматомикозов — это грибы различных родов, основными из которых являются Тrichophyton, Microsporum и Epidermophyton. Также выделяют грибы рода Candida и недерматофитные плесневые грибы, присутствие которых возможно только в условиях выраженного иммунодецифита.
Возбудители проникают в толщу эпидермиса, могут поражать волосяные фолликулы, повреждать структуру волоса и ногтей. Наиболее частые проявления заболевания ограничиваются поражением наружных слоев кожи, ногтей, и носят название дерматофитии. Более глубокое внедрение возможно при поражении грибами рода Candida и недерматофитными представителями.
Все дерматомикозы являются высокозаразными заболеваниями, так как грибы способны распространяться контактным путем не только при прямом контакте, но и при использовании общих предметов обихода, во время посещения общественных мест с высокой влажностью и риском образования грибковых загрязнений поверхностей — например, в тренажерных залах, банях и саунах всегда отмечается высокий риск заражения грибковой кожной инфекцией.
Инфицирование грибком кожи и ее придатков обычно сопровождается наличием жалоб на зуд, изменение цвета. В ходе осмотра обычно можно предположить наличие грибкового поражения, но подтвердить инфекцию можно только при помощи лабораторной диагностики. Исследование волос, чешуек кожи, ногтей помогает с точностью определить возбудителя, исключив при этом иные заболевания со схожими проявлениями (псориаз, опоясывающий лишай, эритема).
Исследование ногтевых пластинок или чешуек кожи на патогенные грибы относится к реакции прямой микроскопии. Полученный материал проходит обработку щелочным раствором (гидроксид калия), после чего производится оценка образцов под микроскопом. При грибковом поражении становятся доступны визуализации споры, гифы и псевдогифы. Зная особенности строения и формы различных родов грибов, можно предположить возбудителя. Так, например, грибы рода Candida не образуют истинных спор, зато имеют форму в виде вытянутой дрожжевой клетки, в том время как дерматофиты обычно имеют трубчатую форум и способны существовать в виде спор.
При проведении исследования врачи-лаборанты уделяют особое внимание точной локализации полученного возбудителя. Так, большинство дерматофитов являются нормальной и даже обязательной микрофлорой нашего организма, постоянно пребывая на поверхности кожи. Но при этом они не способны вызывать жалобы, так как их жизнедеятельность является привычной для организма и не приводит к развитию заболевания. Но в случае повышения количества, занесения грибковой флоры в нетипичную для них среду, например, в подногтевое пространство, они из разряда условно-патогенной флоры переходят в разряд патогенов, вызвавших заболевание.
Изучение микроскопического состава предоставленного материала помогает определить наличие грибкового поражения и предположить род грибов. Но для более точной диагностики с определением рода и чувствительности антимикотиков проводят посев полученной материала на флору.
Патогенные грибы » ФГБУ Саратовская МВЛ
Отделом бактериологии, паразитологии и микологии при исследовании трупов индюшат были обнаружены в воздухоносных мешках патогенные грибы-мукор (лат. Mucor) и пеннициллиум (лат.Penicillium). Мукор (лат. Mucor) — род низших плесневых грибов класса зигомицетов , который включает около 60-ти видов. Они широко распространены в верхнем слое почвы, также развиваются на продуктах питания и органических остатках. Некоторые виды вызывают болезни (мукоромикозы) животных и человека, другие используются для получения антибиотиков или в качестве закваски. Колонии гриба, как правило, бежевого или серого цвета, быстро растут (до нескольких сантиметров в высоту). Старые колонии более тёмные из-за образования многочисленных спорангиев со спорами.
Мукормикоз — заболевание хроническое, сопровождающееся развитием в различных органах грануломатозных процессов или язв, некрозов и геморрагических инфарктов. Микоз развивается чаще всего при ослаблении общей резистентности организма. Заражение происходит через дыхательные пути или пищеварительный тракт. Возникновению заболевания способствуют травматические повреждения кожи или слизистых оболочек. Диагноз обычно подтверждают после вскрытия и проведения микологических исследований.
Пенициллиум, кистевик (Penicillium), род несовершенных грибов, сумчатая стадия которых относится к порядку плектасковых. Известно около 250 видов, широко распространённых в почве. Именно эти грибы первыми утилизируют лесные отходы. Пенициллы — аэробы, поверхность зрелых колоний мучнистая благодаря массе зрелых конидий. Оттенки культур самые разнообразные – от беловато-желтых до фиолетовых и темно-коричневых.
Представители рода Penicillium вызывают незаразное заболевание животных и птиц — пенициллиомикоз, которое возникает на фоне снижения общей резистентности организма. Пенициллы поражают кожу, слизистые оболочки, часто выделяются из легких, воздухоносных мешков. Диагностика данного заболевания складывается из исследования патологического материала, выделения чистой культуры и гистологического исследования.
Пенициллы поражают кожу, слизистые оболочки, часто выделяются из легких, воздухоносных мешков. Диагностика данного заболевания складывается из исследования патологического материала, выделения чистой культуры и гистологического исследования.
Имея повсеместное распространение и поражая почти все виды животных и птиц, микозы наносят существенный экономический ущерб. Животные и птицы отстают в росте и развитии. Большие средства расходуются на проведение лечебных и дезинфекционных мероприятий. Нередко данные заболевания заканчиваются летальным исходом, особенно высокая смертность отмечается у птицы.
Мукор под микроскопом. Пенициллиум на питательной среде.
грибов | Микробиология
Цели обучения
- Объясните, почему изучение грибов, таких как дрожжи и плесень, относится к дисциплине микробиологии
- Опишите уникальные характеристики грибов
- Описать примеры бесполого и полового размножения грибов
- Сравните основные группы грибов в этой главе и приведите примеры каждой
- Определите примеры основных причин инфекций, вызываемых дрожжами и плесенью
- Определите примеры токсин-продуцирующих грибов
- Классифицировать грибковые организмы по основным группам
Грибы включают разнообразную группу организмов, которые являются гетеротрофными и обычно сапрозойскими.Помимо хорошо известных макроскопических грибов (таких как грибы и плесень), микроскопическими являются многие одноклеточные дрожжи и споры макроскопических грибов. По этой причине грибы включены в область микробиологии.
Грибы важны для человека по-разному. Как микроскопические, так и макроскопические грибы имеют медицинское значение, при этом некоторые патогенные виды могут вызывать микозы и (болезни, вызываемые грибами). Некоторые патогенные грибы являются условно-патогенными, что означает, что они в основном вызывают инфекции, когда иммунная защита хозяина нарушена, и обычно не вызывают болезни у здоровых людей. Грибы важны и в других отношениях. Они действуют как разлагатели в окружающей среде и имеют решающее значение для производства определенных продуктов, таких как сыры. Грибы также являются основными источниками антибиотиков, таких как пенициллин из грибка Penicillium .
Грибы важны и в других отношениях. Они действуют как разлагатели в окружающей среде и имеют решающее значение для производства определенных продуктов, таких как сыры. Грибы также являются основными источниками антибиотиков, таких как пенициллин из грибка Penicillium .
Характеристики грибов
Грибы обладают четко выраженными характеристиками, которые отличают их от других организмов. Большинство многоклеточных грибковых тел, обычно называемых плесневыми грибами , состоят из нитей, называемых гифами .Гифы могут образовывать запутанную сеть, называемую мицелием , и образовывать слоевище (тело) мясистых грибов. Гифы, у которых есть стенки между клетками, называются перегородочными гифами ; гифы, у которых отсутствуют стенки и клеточные мембраны между клетками, называются несептатными или ценоцитарными гифами () (Рисунок 1).
Рис. 1. Многоклеточные грибы (плесневые грибки) образуют гифы, которые могут быть септатными или несептированными. Клетки одноклеточных грибов (дрожжей) образуют псевдогифы из отдельных дрожжевых клеток.
В отличие от плесени, дрожжи — одноклеточные грибы. Почкующиеся дрожжи размножаются бесполым путем, отпочковываясь от более мелкой дочерней клетки; полученные клетки могут иногда слипаться в виде короткой цепи или псевдогифа (Рисунок 1). Candida albicans — обычные дрожжи, образующие псевдогифы; он связан с различными инфекциями у людей, включая вагинальные дрожжевые инфекции, кандидоз полости рта и кандидоз кожи.
Некоторые грибы диморфны и имеют более одного вида в течение своего жизненного цикла.Эти диморфные грибы могут проявляться как дрожжи или плесень, что может иметь важное значение для инфекционности. Они способны изменять свой внешний вид в ответ на изменения окружающей среды, такие как доступность питательных веществ или колебания температуры, расти как плесень, например, при 25 ° C (77 ° F), и как дрожжевые клетки при 37 ° C (98,6 ° F). F). Эта способность помогает диморфным грибам выживать в различных средах. Histoplasma capsulatum , патоген, вызывающий гистоплазмоз , инфекцию легких, является примером диморфного гриба (рис. 2).
F). Эта способность помогает диморфным грибам выживать в различных средах. Histoplasma capsulatum , патоген, вызывающий гистоплазмоз , инфекцию легких, является примером диморфного гриба (рис. 2).
Рисунок 2. Histoplasma capsulatum — диморфный гриб, который растет в почве, подверженной воздействию фекалий птиц или летучих мышей (гуано) (вверху слева). Он может менять форму, чтобы выжить при разных температурах. На открытом воздухе он обычно растет как мицелий (как показано на микрофотографии, внизу слева), но при вдыхании спор (справа) он реагирует на высокую внутреннюю температуру тела (37 ° C [98,6 ° F] ), превращаясь в дрожжи, которые могут размножаться в легких, вызывая гистоплазмоз хронического заболевания легких.(кредит: модификация работы Центров по контролю и профилактике заболеваний)
Стенки и мембраны грибковых клеток обладают уникальными особенностями. Стенки грибковых клеток содержат хитин , в отличие от целлюлозы, обнаруженной в клеточных стенках растений и многих простейших. Кроме того, в то время как животные содержат холестерин в клеточных мембранах, мембраны грибковых клеток содержат разные стерины, называемые эргостеролами . Эргостерины часто используются как мишени для противогрибковых препаратов.
Жизненные циклы грибов уникальны и сложны. Грибы размножаются половым путем либо путем перекрестного, либо самооплодотворения. Гаплоидные грибы образуют гифы с гаметами на концах. Используются два разных типа спаривания (представленные как «+ тип» и «- тип»). Цитоплазмы гамет + и — сливаются (событие, называемое плазмогамией), образуя клетку с двумя различными ядрами (дикариотическая клетка ). Позже ядра сливаются (событие называется кариогамия ), образуя диплоидную зиготу.Зигота подвергается мейозу с образованием спор , которые прорастают, чтобы начать гаплоидную стадию, которая в конечном итоге создает больше гаплоидного мицелия (Рисунок 3). В зависимости от таксономической группы эти споры, образующиеся половым путем, известны как зигоспоры (у Zygomycota), аскоспоры (у Ascomycota) или базидиоспоры (у Basidiomycota) (рис. 4).
В зависимости от таксономической группы эти споры, образующиеся половым путем, известны как зигоспоры (у Zygomycota), аскоспоры (у Ascomycota) или базидиоспоры (у Basidiomycota) (рис. 4).
Грибы также могут демонстрировать бесполое размножение митозом, митоз с почкованием, фрагментацию гиф и образование бесполых спор митозом.Эти споры представляют собой специализированные клетки, которые, в зависимости от организма, могут обладать уникальными характеристиками для выживания, размножения и распространения. У грибов есть несколько типов бесполых спор, которые могут иметь важное значение при классификации.
Рисунок 3. Щелкните, чтобы увеличить изображение. У зигомицетов есть половой и бесполый жизненные циклы. В половом жизненном цикле типы спаривания + и — спрягаются, образуя зигоспорангий.
Подумай об этом
- Диморфный гриб — это дрожжи или плесень? Объяснять.
Разнообразие грибов
Грибы очень разнообразны и состоят из семи основных групп. Не все из семи групп содержат патогены. Некоторые из этих групп обычно связаны с растениями и включают патогены растений. Например, урединиомицеты и устилагомицеты включают растение , ржавчину, и головню, , соответственно. Они образуют красноватые или темные массы на растениях соответственно в виде ржавчины (красный цвет) или головни (темный цвет). Некоторые виды имеют существенное экономическое влияние из-за их способности снижать урожайность сельскохозяйственных культур.Glomeromycota включает микоризные грибы , важные симбионты с корнями растений, которые могут способствовать росту растений, действуя как расширенная корневая система. Glomeromycota — облигатные симбионты, а это означает, что они могут выжить, только если связаны с корнями растений; грибы получают углеводы из растений, а растения получают выгоду от повышенной способности поглощать питательные вещества и минералы из почвы. Chytridiomycetes ( chytrids ) — небольшие грибы, но они чрезвычайно важны с экологической точки зрения. Хитриды обычно водные и имеют жгутиковые подвижные гаметы; определенные типы причастны к сокращению количества земноводных по всему миру. Из-за их медицинской важности мы сосредоточимся на Zygomycota, Ascomycota, Basidiomycota и Microsporidia. На рисунке 9 приведены характеристики этих важных с медицинской точки зрения групп грибов.
Хитриды обычно водные и имеют жгутиковые подвижные гаметы; определенные типы причастны к сокращению количества земноводных по всему миру. Из-за их медицинской важности мы сосредоточимся на Zygomycota, Ascomycota, Basidiomycota и Microsporidia. На рисунке 9 приведены характеристики этих важных с медицинской точки зрения групп грибов.
Zygomycota (зигомицеты) — это в основном сапрофиты с ценоцитарными гифами и гаплоидными ядрами. Они используют спорангиоспоры для бесполого размножения. Название группы происходит от зигоспор , которые они используют для полового размножения (рис. 3), у которых есть твердые стенки, сформированные в результате слияния репродуктивных клеток двух особей.Зигомицеты важны для науки о продуктах питания и как патогены сельскохозяйственных культур. Одним из примеров является Rhizopus stolonifer (рис. 4), важная хлебная плесень, которая также вызывает ожог проростков риса. Mucor — это род грибов, которые потенциально могут вызывать некротические инфекции у людей, хотя большинство видов не переносят температуры, обнаруживаемые в телах млекопитающих (рис. 4).
Рис. 4. На этих изображениях показаны споры, полученные бесполым путем. (а) На этой микрофотографии в светлом поле показано высвобождение спор из спорангия на конце гифы, называемого спорангиофором.Организм — это Mucor sp. грибок, плесень, часто встречающаяся в помещении. (b) Спорангии растут на концах стеблей, которые выглядят как белый пух, видимый на этой хлебной форме, Rhizopus stolonifer . Кончики хлебной плесени представляют собой темные спорангии, содержащие споры. (кредит а: модификация работы Центров по контролю и профилактике заболеваний; кредит б справа: модификация работы «Эндрю» / Flickr)
Ascomycota включает грибы, которые используются в пищу (съедобные грибы, сморчки и трюфели), другие грибы, которые являются частыми причинами порчи пищевых продуктов (хлебные плесени и патогенные микроорганизмы растений), а также другие, которые являются патогенами человека. У Ascomycota могут быть перегородчатые гифы и чашевидные плодовые тела, называемые ascocarps . Некоторые роды Ascomycota используют аскоспоры , образующиеся половым путем, , а также бесполые споры, называемые конидиями , но половые фазы для других не обнаружены и не описаны. Некоторые производят аск , содержащий аскоспоры внутри аскокарпа (рис. 5).
У Ascomycota могут быть перегородчатые гифы и чашевидные плодовые тела, называемые ascocarps . Некоторые роды Ascomycota используют аскоспоры , образующиеся половым путем, , а также бесполые споры, называемые конидиями , но половые фазы для других не обнаружены и не описаны. Некоторые производят аск , содержащий аскоспоры внутри аскокарпа (рис. 5).
Рис. 5. (a) На этой микрофотографии в светлом поле показаны аскоспоры, высвобождающиеся из асков у гриба Talaromyces flavus var. flavus . (b) На этой электронной микрофотографии показаны конидии (споры), переносимые конидиофором Aspergillus , типом токсичного гриба, встречающегося в основном в почве и растениях. (c) На этой микрофотографии в светлом поле показаны дрожжи Candida albicans , возбудитель кандидоза и молочницы. (кредит a, b, c: модификация работы Центров по контролю и профилактике заболеваний)
Рис. 6. Эти аскоспоры, выстроенные внутри аска, образуются половым путем. (Источник: Питер Г.Вернер)
Примеры Ascomycota включают несколько форм для хлеба и второстепенных патогенов, а также виды, способные вызывать более серьезные микозы. Виды рода Aspergillus являются важными причинами аллергии и инфекций и используются в исследованиях и при производстве некоторых ферментированных алкогольных напитков, таких как японское саке . Грибок Aspergillus flavus , загрязнитель орехов и хранимых зерен, продуцирует афлатоксин , который является одновременно токсином и наиболее мощным из известных природных канцерогенов. Neurospora crassa особенно полезна в генетических исследованиях, потому что споры, образующиеся в результате мейоза, хранятся внутри аска в ряду, который отражает клеточные деления, которые их произвели, что дает прямое представление о сегрегации и ассортименте генов (рис. 6). . Penicillium производит антибиотик пенициллин (рис. 5).
5).
Многие виды аскомицетов важны с медицинской точки зрения. Большое количество видов в родах: Trichophyton , Microsporum и Epidermophyton — это дерматофиты , патогенные грибы, способные вызывать кожные инфекции, такие как микоз стопы, зуд спортсмена и стригущий лишай . Blastomyces dermatitidis — диморфный гриб, вызывающий бластомикоз , респираторную инфекцию, которая, если ее не лечить, может распространиться на другие участки тела, иногда приводя к смерти. Другой важный респираторный патоген — диморфный гриб Histoplasma capsulatum (рис. 2), , , который ассоциируется с птицами и летучими мышами в долинах рек Огайо и Миссисипи. Coccidioides immitis вызывает серьезное заболевание легких Лихорадка долины . Candida albicans , наиболее частая причина вагинальных и других дрожжевых инфекций, также является аскомицетным грибком; он является частью нормальной микробиоты кожи, кишечника, половых путей и ушей (рис. 5). Аскомицеты также вызывают болезни растений, в том числе инфекции спорыньи, болезнь голландского вяза и мучнистую росу.
Saccharomyces дрожжи, включая пекарские дрожжи S. cerevisiae , являются одноклеточными аскомицетами с гаплоидной и диплоидной стадиями (рис. 7).Этот и другие видов сахаромицетов используются для пивоварения.
Рисунок 7. Щелкните, чтобы увеличить изображение. Жизненный цикл аскомицета характеризуется образованием аски во время половой фазы. Гаплоидная фаза — преобладающая фаза жизненного цикла.
Basidiomycota (базидиомицеты) — это грибы, которые имеют базидий (булавовидные структуры), которые производят базидиоспоры (споры, образующиеся в результате бутонизации) внутри плодовых тел, называемых базидиокарпами (рис. 8).Они важны как разлагатели и как пища. В эту группу входят ржавчина, вонючка, лепешки и грибы. Особое значение имеют несколько видов. Cryptococcus neoformans , грибок, обычно встречающийся в окружающей среде в виде дрожжей, может вызывать серьезные легочные инфекции при вдыхании людьми с ослабленной иммунной системой. Съедобный луговой гриб Agricus campestris является базидиомицетом, как и ядовитый гриб Amanita phalloides , известный как «смертельная шапка».Смертельные токсины, продуцируемые A. phalloides , были использованы для изучения транскрипции.
8).Они важны как разлагатели и как пища. В эту группу входят ржавчина, вонючка, лепешки и грибы. Особое значение имеют несколько видов. Cryptococcus neoformans , грибок, обычно встречающийся в окружающей среде в виде дрожжей, может вызывать серьезные легочные инфекции при вдыхании людьми с ослабленной иммунной системой. Съедобный луговой гриб Agricus campestris является базидиомицетом, как и ядовитый гриб Amanita phalloides , известный как «смертельная шапка».Смертельные токсины, продуцируемые A. phalloides , были использованы для изучения транскрипции.
Рисунок 8. Щелкните, чтобы увеличить изображение. Жизненный цикл базидиомицета чередует гаплоидное поколение с продолжительной стадией, на которой в гифах присутствуют два ядра (дикарион).
Наконец, Microsporidia — одноклеточные грибы, которые являются облигатными внутриклеточными паразитами. У них отсутствуют митохондрии, пероксисомы и центриоли, но их споры выделяют уникальный полярный каналец , который проникает через мембрану клетки-хозяина, позволяя грибку проникнуть в клетку.Ряд микроспоридий являются патогенами человека, и инфекции, вызываемые микроспоридиями, называются микроспоридиозом . Одним из патогенных видов является Enterocystozoan bieneusi , который может вызывать такие симптомы, как диарея, холецистит (воспаление желчного пузыря) и, в редких случаях, респираторные заболевания.
| Таблица 1. Выберите группы грибов | ||||
|---|---|---|---|---|
| Группа | Характеристики | Примеры | Виды, важные с медицинской точки зрения | Изображение |
| Аскомикота | Перегородчатые гифы Аскус с аскоспорами в аскокарпе Конидиоспоры | Чашка грибов Съедобные грибы Сморчки Трюфели Нейроспора Пенициллим | Aspergillus spp. Trichophyton spp. Microsporum spp. Epidemophyton spp. Blastomyces demititidis Histoplasma capsulatum | Aspergillus niger |
| Basidiomycota | Базидия Образует базидиоспоры в базидиокарпии | Клуб грибов Рустов Стинхорс Снежинки Грибы Криптококк neoformans Мухомор фаллоидный | Криптококк neoformans | Мухомор фаллоидный |
| Microsporidia | Отсутствие митохондрий, пероксисом и центриолей Споры образуют полярную трубку | Enterocystozoan bieneusi | Enterocystozoan bieneusi | Microsporidia (не идентифицировано) |
| Zygomycota | В основном сапрофиты Ценоцитарные гифы Гаплоидные ядра Зигоспоры | Rhizopus stolonifera | Mucor spp. | Rhizopus sp. |

Подумай об этом
- Какая группа грибов вызывает наибольшее количество болезней человека?
Эукариотические патогены в эукариотических хозяевах
Когда мы думаем об антимикробных препаратах, на ум часто приходят такие антибиотики, как пенициллин. Пенициллин и родственные антибиотики препятствуют синтезу клеточных стенок пептидогликана, который эффективно воздействует на бактериальные клетки.Эти антибиотики полезны, потому что люди (как и все эукариоты) не имеют клеточных стенок пептидогликана.
Сложнее разработать лекарства, которые эффективны против эукариотических клеток, но не вредны для клеток человека. Несмотря на огромные морфологические различия, клетки людей, грибов и простейших схожи по своим рибосомам, цитоскелетам и клеточным мембранам. В результате становится все сложнее разработать лекарства, которые нацелены на простейшие и грибы так же, как антибиотики на прокариот.
В результате становится все сложнее разработать лекарства, которые нацелены на простейшие и грибы так же, как антибиотики на прокариот.
Фунгициды имеют относительно ограниченные механизмы действия. Поскольку в клеточных мембранах грибов содержится эргостерин (вместо холестерина), различные ферменты, участвующие в производстве стеролов, могут быть мишенью для некоторых лекарств. Фунгициды азол и морфолин препятствуют синтезу мембранных стеринов. Они широко используются в сельском хозяйстве (фенпропиморф) и в клинической практике (например, миконазол). Некоторые противогрибковые препараты нацелены на клеточные стенки хитина грибов.Несмотря на успех этих соединений в борьбе с грибами, противогрибковые препараты для системных инфекций по-прежнему имеют более токсичные побочные эффекты, чем антибиотики для бактерий.
Клиническая направленность: Энтони, часть 3
Этот пример продолжает историю Энтони, начатую в книге «Одноклеточные эукариотические паразиты и паразитические гельминты».
Энтони чувствует облегчение, что стригущий лишай — не настоящий червь, но хочет знать, что это такое на самом деле. Врач объясняет, что стригущий лишай — это грибок.Он говорит Энтони, что не увидит, как грибы вылезают из его кожи, потому что этот гриб больше похож на невидимую часть гриба, которая прячется в почве. Их врач заверяет Энтони, что они собираются вылечить и от него грибок.
Врач очищает, а затем осторожно соскабливает поражение, чтобы поместить образец на предметное стекло. Глядя на это под микроскопом, врач может подтвердить, что поражение Энтони вызвано грибковой инфекцией. На рисунке 9 можно увидеть макро- и микроконидии в Trichophyton rubrum .Также видны клеточные стенки. Даже если бы возбудитель походил на гельминта под микроскопом, наличие клеточных стенок исключило бы возможность, потому что клетки животных не имеют клеточных стенок.
Рис. 9. На этой микрофотографии показаны гифы (макроконидии) и микроконидии Trichophyton rubrum, дерматофита, ответственного за грибковые инфекции кожи. (кредит: модификация работы Центров по контролю и профилактике заболеваний)
Врач прописывает матери Энтони противогрибковый крем для лечения стригущего лишая.Мать Энтони спрашивает: «Что нам делать, если он не проходит?»
- Можно ли лечить все формы стригущего лишая с помощью одного и того же противогрибкового препарата?
Мы вернемся к примеру Энтони на следующих страницах.
Ключевые концепции и резюме
- Грибы включают разнообразные сапротрофные эукариотические организмы с хитиновыми клеточными стенками
- Грибы могут быть одноклеточными и многоклеточными; некоторые (например, дрожжи) и споры грибов микроскопические, а некоторые крупные и заметные
- Репродуктивные типы важны для различения грибковых групп
- Важные с медицинской точки зрения виды существуют в четырех группах грибов: Zygomycota, Ascomycota, Basidiomycota и Microsporidia
- Члены Zygomycota, Ascomycota и Basidiomycota производят смертельные токсины
- Важные различия в грибковых клетках, такие как эргостерины в грибковых мембранах, могут быть мишенями для противогрибковых препаратов, но сходство между человеческими и грибковыми клетками затрудняет поиск мишеней для лекарств, и эти лекарства часто имеют токсические побочные эффекты
.
Множественный выбор
К каким из следующих грибов относятся?
- конидии
- ascus
- полярный каналец
- базидиокарпий
Показать ответ
Ответ d.Грибы — это базидиокарпий.
Что из перечисленного является наиболее частой причиной дрожжевых инфекций у человека?
- Candida albicans
- Blastomyces dermatitidis
- Криптококк neoformans
- Aspergillus fumigatus
Показать ответ
Ответ а. Candida albicans — наиболее частая причина дрожжевых инфекций человека.
Candida albicans — наиболее частая причина дрожжевых инфекций человека.
Что из перечисленного представляет собой грибок аскомицета, связанный с пометом летучих мышей, который может вызвать респираторную инфекцию при вдыхании?
- Candida albicans
- Histoplasma capsulatum
- Rhizopus stolonifera
- Trichophyton rubrum
Показать ответ
Ответ б. Histoplasma capsulatum — грибок аскомицет, связанный с пометом летучих мышей.
Заполните бланк
Гифы без перегородки также называются _________.
Покажи ответ
Несептатные гифы также называют ценоцитами .
Одноклеточные грибы называются _________.
Покажи ответ
Одноклеточные грибы называются дрожжами .
Некоторые грибы оказались полезными с медицинской точки зрения, поскольку их можно использовать для производства _________.
Покажи ответ
Некоторые грибы оказались полезными с медицинской точки зрения, поскольку их можно использовать для производства антибиотиков .
Подумай об этом
- Какие роды грибов являются распространенными дерматофитами (грибами, вызывающими кожные инфекции)?
- Что такое дикариотическая клетка?
- Объясните преимущества исследований путей, участвующих в синтезе хитина в грибах.
- На каком из рисунков изображены перегородчатые гифы?
Термодиморфные грибковые патогены человека — полифилетические патогены с конвергентным признаком патогенности
Cold Spring Harb Perspect Med.2015 Авг; 5 (8): a019794.
1 и 2
Анита Сил
1 Кафедра микробиологии и иммунологии Калифорнийского университета, Сан-Франциско, Калифорния 94143
Алекс Андрианопулос
2 Департамент генетики Мельбурнского университета , Виктория 3010, Австралия
1 Кафедра микробиологии и иммунологии Калифорнийского университета, Сан-Франциско, Калифорния 94143
2 Кафедра генетики Мельбурнского университета, Виктория 3010, Австралия
Автор, отвечающий за переписку. Авторское право © 2015 Лаборатория Колд Спринг Харбор Пресс; все права защищеныЭта статья цитировалась в других статьях в PMC.
Авторское право © 2015 Лаборатория Колд Спринг Харбор Пресс; все права защищеныЭта статья цитировалась в других статьях в PMC.
Abstract
Грибы умеют изменять форму своих клеток и программу развития в ответ на сигналы в их окружении. Здесь мы сосредоточимся на группе эволюционно связанных грибковых патогенов человека, известных как термически диморфные грибы. Эти организмы растут в форме гиф в окружающей среде, но резко меняют свою морфологию в организме-хозяине-млекопитающем.Температура является одним из основных сигналов хозяина, который инициирует их преобразование в форму «хозяина», и его достаточно в лаборатории для запуска этой адаптированной к хозяину программы развития. Здесь мы обсуждаем основные патогены человека в этой группе: Blastomyces dermatiditis , Coccidioides immitis / posadasii , Histoplasma capsulatum , Paracoccidioides brasiliensis / lutzii (ранее известный как Penicillium marneffei ).Большинство этих организмов являются первичными патогенами, способными вызывать заболевание у здоровых людей, встречающихся с ними в эндемичных районах.
Диморфизм определяется как способность гриба генерировать свободноживущие вегетативные типы клеток, которые являются дрожжевыми или гифальными (), хотя мы обсуждаем некоторые исключения из этого точного определения ниже. Самое экономное объяснение происхождения грибов начинается с одноклеточной эукариотической клетки, из которой произошли многоклеточные нитчатые формы.Предположительно, первые нитчатые формы были бы заблокированы этим способом роста с помощью простых мутаций в аппарате деления клеток, и со временем эволюционировал регулируемый переход между одноклеточными и многоклеточными вегетативными формами (диморфизм). Для нитчатой формы роста способность генерировать независимые одноклеточные формы возникала несколько раз под видом современных программ развития, таких как бесполое и половое размножение, которые производят дифференцированные спящие споры.
Упрощенная схема терморегулирующих форм термодиморфных грибковых возбудителей. Показаны простые изображения окружающей среды и форм хозяев каждого вида грибов. В форме окружающей гифы этих организмов овальные вздутия изображают вегетативные конидии. В случае H. capsulatum показаны туберкулезные макроконидии и микроконидии. В случае Coccidioides spp. Показаны разъединяющие артроконидии. Показаны некоторые характерные различия между фазой роста дрожжей: Blastomyces dermatitidis дрожжевые клетки имеют широкую шейку почки, Paracoccidioides дрожжи могут быть многопучковыми, а T.marneffei дрожжи делятся не бутонированием, а делением. Обратите внимание, что относительный масштаб различных типов клеток (внутри и между видами) не является точным. (Иллюстрация предоставлена Давиной Хокинг Мюррей.)
Диморфизм у грибов, вероятно, возникал независимо несколько раз. Полифилетическая природа диморфизма проявляется в распределении организмов с этой способностью по царству грибов (см.). Диморфные грибы существуют в трех основных типах грибов: Ascomycota, Basidiomycota и Zygomycota.У Ascomycota они распределены по нескольким отрядам и тесно связаны со многими недиморфными грибами. Самый крупный кластер термодиморфных грибов включает Histoplasma capsulatum , Blastomyces dermatiditis , Coccidioides и Paracoccidioides вида отряда Onygenales. Точно так же существует ряд диморфных видов в отряде Ophiostomatales, включая различные виды Sporothrix . Напротив, Talaromyces marneffei — единственный известный диморфный вид в большом отряде Eurotiales и фактически единственный диморфный гриб, у которого дрожжевые клетки делятся путем деления, а не почкования.В других типах известно гораздо меньше известных диморфных грибов, наиболее известными из которых являются базидиомицеты — кукурузная головня Ustilago maydis и зигомицеты — патогены животных, представленные различными видами Mucor . Действительно, многие диморфные грибы являются патогенами животных или растений.
Действительно, многие диморфные грибы являются патогенами животных или растений.
Молекулярно-филогенетический анализ диморфных грибковых патогенов. Древо эволюции основано на сравнении белков основных диморфных патогенов человека и включает в себя ряд других грибковых патогенов для справки.Человеческие патогены (красный шрифт), патогены растений (зеленый шрифт) и непатогены (черный шрифт) показаны и помечены как истинные диморфные виды со свободноживущими типами вегетативных клеток (синий кружок) и виды с морфологическими переходами, которые не являются свободноживущими типы вегетативных клеток (синий треугольник). За исключением C. neoformans и U. maydis , которые принадлежат к типу Basidiomycota , все остальные виды принадлежат к типу Ascomycota .Эти виды охватывают широкий диапазон отрядов ( Onygenales , зеленый; Eurotiales, , желтый; Ophiostomales, , оранжевый; Magnaporthales, коричневый; и Saccharomycetales, , красный). Для создания филогенетических взаимоотношений между организмами домен Pfam Gcd10p был использован для идентификации последовательностей Gcd10 от 16 представляющих интерес видов. CLUSTALX 2.1 (Larkin et al. 2007) использовался для выравнивания полноразмерных белковых последовательностей и генерации самонастраиваемого дерева соединения соседей (1000 бутстрепов; все внутренние узлы имели значение начальной загрузки не менее 700).(Марк Вурхис внес свой вклад в этот анализ.)
Как упоминалось выше, диморфизм в самом строгом смысле включает способность гриба генерировать два типа вегетативных клеток — дрожжевые или гифальные по морфологии. Coccidioides виды не совсем подходят под это определение, потому что они производят не свободноживущие вегетативные дрожжевые клетки, а скорее сферулы, которые охватывают многочисленные одиночные клетки, известные как эндоспоры (). Ряд других грибов демонстрируют аспекты диморфизма в паттернах роста и морфогенеза.Хорошо охарактеризованным примером является рост псевдогифала в аскомицетных дрожжах Saccharomyces cerevisiae ; когда диплоидные клетки S. cerevisiae испытывают ограничение по азоту, они претерпевают морфологический переход к удлиненной клеточной форме с униполярным почкованием и неполным разделением материнских и дочерних клеток. Это приводит к образованию гифоподобных нитей и является программой ряда дрожжей, включая патоген человека Candida albicans , который может образовывать как псевдогифы, так и настоящие гифы.Базидиомицетные дрожжи Cryptococcus neoformans также способны образовывать гифальные волокна в рамках программы спаривания.
Как описано ниже, термически диморфные грибковые патогены являются эндемичными во всем мире. Тем не менее, наше понимание экологической ниши и эпидемиологии этих диморфных патогенов далеко не полное. Поскольку эти организмы реплицируются в отсутствие хозяина-млекопитающего, давление окружающей среды, которое отобрано для специфических для хозяина признаков, неясно.Все эти грибы демонстрируют устойчивый, хотя иногда и медленный, прототрофный рост в форме гиф в окружающей среде. Эти гифы образуют бесполые споры (конидии), которые часто представляют собой наиболее распространенный инфекционный возбудитель для данного организма. Считается, что конидиальные гифы существуют в смеси неуловимых сред, не являющихся хозяевами, и роста на мертвых хозяевах. Интересно, что было высказано предположение, что эукариотические хищники, такие как амебы и слизистые плесени, могут оказывать селективное давление на признаки вирулентности, которые необходимы для выживания антимикробного натиска фагоцитарных клеток млекопитающих (Steenbergen et al.2004 г.).
Blastomyces dermatitidis
Первое известное сообщение о заражении B. dermatitidis произошло в 1894 году, когда T.C. Гилкрист сообщил о пациенте с кожным заболеванием, предположительно вызванным простейшей инфекцией. Однако Гилкрист и его коллеги вскоре заметили зарождающуюся дрожжевую форму в тканях и культивировали организм гиф из образцов пациентов, которые они назвали B. dermatitidis (Gilchrist 1894; Gilchrist and Stokes 1896, 1898). После открытия Гилкриста потребовалось еще как минимум 50 лет, чтобы понять, что кожные проявления заболевания связаны с поражением легких, что позволяет предположить, что все случаи поражения кожи являются результатом распространения из легких.Теперь ясно, что бластомикоз — это в первую очередь инфекция легких, которая может распространяться на кожу, костно-суставные структуры, мочеполовые пути и другие органы (Smith and Kauffman 2010).
B. dermatitidis сложно изолировать от окружающей среды, поэтому эпидемиология бластомикоза менее определена, чем гистоплазмоз или кокцидиомикоз. Основываясь в основном на сообщениях о случаях заболевания, B. dermatitidis , как известно, является эндемиком в долинах рек Миссисипи и Огайо, штатах Среднего Запада, провинциях Канады, граничащих с Великими озерами, а также в районе Нью-Йорка и Канады, прилегающего к реке Св.Лоуренс Сивей (Кляйн и др. 1986, 1987; Крэмптон и др. 2002; Кано и др. 2003; Дворкин и др. 2005). Кроме того, во всем мире зарегистрированы спорадические случаи бластомикоза (De Groote et al. 2000; Arnett et al. 2008; Smith and Kauffman 2010). Большинство случаев протекает бессимптомно, но у людей, которые либо подвергаются воздействию большой дозы инфекционных частиц, либо имеют дефект клеточного иммунитета, развивается более тяжелое прогрессирующее заболевание (Maresca and Kobayashi 2000; Smith and Kauffman 2010).В редких случаях может развиться острый респираторный дистресс-синдром (Smith and Kauffman 2010), но хронические поражения кожи и костей являются наиболее частыми внелегочными осложнениями. Заболеваемость у мужчин выше, чем у женщин. Кроме того, собаки в эндемичных регионах подвержены бластомикозу, что делает его важной ветеринарной проблемой (Baumgardner et al. 1995).
B. dermatitidis претерпевает температурно-зависимый морфогенез, вырастая в форме гиф в лаборатории при температуре ниже 30 ° C и в дрожжевой форме при 37 ° C () (Maresca and Kobayashi 2000; Nemecek et al.2006 г.). Как и у других термодиморфных грибов, его морфология обратима путем изменения температуры окружающей среды. Заражение происходит при вдыхании конидий или фрагментов гиф, а преобразование в дрожжевую форму происходит внутри хозяина. В отличие от H. capsulatum , который обнаруживается внутриклеточно внутри фагоцитов во время инфекции, B. dermatitidis растет как внеклеточные дрожжи в микроабцессах (Maresca and Kobayashi 2000; Smith and Kauffman 2010). B. dermatitidis дрожжи, как правило, имеют широкую шейку между маткой и почкой (), а дрожжевые клетки обычно, но не всегда, больше, чем у H.capsulatum .
Ряд фундаментальных инструментов позволил идентифицировать факторы вирулентности и изучить основные процессы в B. dermatitidis , включая идентификацию иммуномодулирующего адгезина Bad1, а также факторов, регулирующих морфогенез и гомеостаз железа (Brandhorst et al. 2002; Sullivan et al.2002; Krajaejun et al.2007; Gauthier et al.2010). Огромным достижением в понимании терморегулируемого морфогенеза стала идентификация гибридной гистидинкиназы, которая необходима для роста дрожжевой фазы и индукции гена вирулентности в ответ на температуру как в B.dermatitidis и H. capsulatum (Nemecek et al. 2006). Кроме того, плодотворные исследования ответа хозяина на Blastomyces и другие термически диморфные грибы дали глубокое понимание иммунобиологии грибковых инфекций (Wuthrich et al. 2011, 2012; Wang et al. 2014).
Coccidioides immitis / posadasii
В 1891 году студент-медик из Аргентины по имени Алехандро Посадас увидел пациента с необычным поражением кожи. Посадас и Роберт Вернике отметили в поражении кокцидоподобного «паразита» (Posadas 1892; Wernicke 1892).Первый случай заболевания в Северной Америке был зарегистрирован в 1894 году (Rixford 1894). В течение следующих 20 лет как диморфизм организма (Ophuls and Moffitt, 1900), так и его распространенность в Калифорнии (Dickson 1915) стали очевидными.
Coccidioides — патоген Нового Света. Род Coccidioides недавно был разделен на два вида (Fisher et al. 2002): immitis , эндемик центральной и южной Калифорнии и Северной Мексики; и posadasii , обнаруженных по всей Аризоне, Техасе, Мексике и в некоторых частях Южной Америки (Chiller et al.2003; Galgiani et al. 2005; Пфаллер и Дикема 2010; Марсден-Хауг и др. 2013). Coccidioides ассоциируется с мелкими млекопитающими в окружающей среде (Ashburn and Emmons 1942; Emmons 1962), хотя точная природа этой ассоциации и ее роль в экологии организма несколько противоречивы.
Заболевание, вызываемое видами Coccidioides , кокцидиоидомикозом, также известно как лихорадка долины Сан-Хоакин или просто лихорадка долины из-за ее распространенности в Центральной долине Калифорнии.Примечательно, что число случаев заражения Coccidioides, растет в эндемичных районах, причем в период с 2001 по 2006 год заболеваемость в Аризоне и Калифорнии увеличилась более чем на 90%. В результате это воздействие на общественное здоровье огромно (Hector et al. 2011).
Coccidioides споры (артроконидии) образуются из чередующихся клеток гиф (). По мере дифференциации гиф отдельные клетки отделяются друг от друга, и чередующиеся клетки становятся бочкообразными артроконидиями 3-5 мкм, в то время как остальные клетки подвергаются автолизу.Попав в организм млекопитающего-хозяина, артроконидии претерпевают резкую трансформацию, в конечном итоге создавая структуру, уникальную для рода Coccidioides (). Артроконидии претерпевают изотропный рост, увеличиваясь в размерах и модифицируя клеточную стенку с образованием сферулы — клеточной структуры до 100 мкм в диаметре. Сферула содержит от 100 до 300 одноклеточных единиц, известных как эндоспоры. Разрыв сферулы в тканях хозяина высвобождает эти эндоспоры, каждая из которых способна развиваться в новую сферулу (Nguyen et al.2013). Сама сферула рефрактерна к фагоцитозу. Формирование сфер можно вызвать in vitro путем культивирования гриба в жидкой модифицированной среде Converse при 37–40 ° C (Converse 1955; Sun et al. 1976).
Как и в случае с большинством грибов, обсуждаемых в этой статье, преобладающий путь заражения Coccidioides spp. это ингаляция. Большинство случаев протекает бессимптомно, но у некоторых пациентов развивается осложненная пневмония. Диссеминированный кокцидиоидомикоз относится к заболеванию за пределами грудной клетки, чаще всего к таким участкам, как кожа, суставы, кости и мозговые оболочки.Реже встречается распространение на гортань, живот, придатки и перикард. У афроамериканцев риск развития диссеминированного кокцидиоидомикоза выше, чем у представителей других этнических групп, что позволяет предположить, что генетический компонент хозяина влияет на тяжесть исхода заболевания (Паппагианис и др., 1979; Паппагианис, 1988; Киркланд и Фирер, 1996; Нгуен и др., 2013). ). Подобно другим термодиморфным грибковым патогенам, люди с дефектным клеточным иммунитетом более восприимчивы к тяжелым заболеваниям.
Надежный молекулярный набор инструментов существует для Coccidioides spp. нарушение гена, трансформация, опосредованная Agrobacterium , и профилирование транскрипции определили ряд факторов, критических для патогенеза, таких как иммуномодулирующий гликопротеин внешней стенки шарика (SOWgp) и уреаза (URE), которая, как считается, способствует повреждению тканей хозяина ( Abuodeh et al.2000; Hung et al.2007, 2012). Исследования экспрессии генов Coccidioides гиф и сферул определяют программу транскрипции различных морфологических состояний этого организма (Johannesson et al.2006; Whiston et al. 2012; Viriyakosol et al. 2013), что даст важные ключи к разгадке молекулярной программы, которая запускается в ответ на сигналы хозяина, такие как температура. Кроме того, исследования популяционной геномики используются с большим эффектом для определения генов, которые могут лежать в основе вирулентности отдельных изолятов Coccidioides (Sharpton et al. 2009; Neafsey et al. 2010).
Histoplasma capsulatum
Гистоплазмоз был впервые описан в 1906 году Сэмюэлем Дарлингом, американским врачом, работающим в больнице зоны канала Анкон в Панаме (Darling 1906).Во время вскрытия пациента с Мартиники, умершего от разрушительной инфекции, Дарлинг обнаружил внутриклеточные организмы, у которых, по-видимому, были капсулы. Он ошибочно определил этот организм как нового простейшего паразита и назвал его H. capsulatum из-за того, что он обитает в «гистиоцитах» или макрофагах. В течение следующих нескольких десятилетий стало очевидно, что H. capsulatum на самом деле является экологическим грибком без капсулы. Этот организм был впервые выделен из почвы Честером Эммонсом в 1948 году (Emmons 1949).
Гистоплазмоз у человека вызывается двумя различными разновидностями H. capsulatum , которые проявляются с очень разными клиническими исходами: разновидностью H. capsulatum capsulatum и разновидностью H. capsulatum разновидностью duboisii (Kauffman 2007). H. capsulatum ( Hc ) вар. capsulatum встречается во всем мире (в основном в Северной, Южной и Центральной Америке, Юго-Восточной Азии и Африке) и ассоциируется с легочными и системными заболеваниями (т.е.э., классический) гистоплазмоз. Напротив, Hc var. duboisii преимущественно встречается в Западной и Центральной Африке. Поскольку Hc var. duboisii вызывает поражения кожи и костей. Первоначально его отличали от сорта capsulatum на основании симптомов заболевания (Cockshott and Lucas 1964). Возбудитель болезни — сорт Hc var. duboisii получил название африканский гистоплазмоз, и на молекулярном уровне об этой разновидности мало что известно.Штаммы разновидностей capsulatum и duboisii были секвенированы Институтом Броуда и Центром секвенирования генома Вашингтонского университета в Сент-Луисе, что позволит пролить свет на фундаментальные молекулярные различия между изолятами H. capsulatum . Кроме того, филогенетический анализ, основанный на вариации геномных последовательностей четырех генов, кодирующих белок, был выполнен на изолятах 137 Histoplasma с шести континентов (Kasuga et al. 2003). Эти изоляты попали в восемь клад, семь из которых представляют разные филогенетические виды.Хотя все Hc var. изолятов duboisii были представлены в африканской кладе, в эту кладу также вошла разновидность Hc var. capsulatum особь. В дополнение к этим двум разновидностям, третья разновидность, Hc var. farciminosum , возбудитель лошадей, был обнаружен в трех кладах. Необходима дополнительная молекулярная работа, чтобы понять биологические различия между этими разновидностями и их связь с болезнями.
Далее мы сосредоточимся на классическом гистоплазмозе, вызванном Hc var. capsulatum , для простоты обозначается как H. capsulatum . Хотя этот организм встречается во всем мире, он, как правило, вызывает наибольшее бремя болезней в Центральной и Северной Америке. Наряду с B. dermatitidis и Coccidioides spp., H. capsulatum считается наиболее частой причиной грибковых инфекций легких у иммунокомпетентных людей.Исследование госпитализаций в США по поводу эндемических микозов показывает, что эти грибы вызывают значительную заболеваемость и смертность у здоровых хозяев в эндемичных регионах (Chu et al., 2006), с примерно 25000 предполагаемых опасных для жизни инфекций в год в Среднем Западе США, вызываемых H . capsulatum инфекция (Brown et al. 2012). В США H. capsulatum является эндемиком в долинах рек Огайо и Миссисипи. Этот организм обнаружен в почве, содержащей большое количество гуано птиц или летучих мышей, и летучие мыши были предложены в качестве вектора распространения, потому что гриб хорошо растет в почве, загрязненной гуано летучих мышей, и потому что сами летучие мыши могут быть колонизированы H.capsulatum (Хофф и Биглер, 1981). Подавляющее большинство человеческих инфекций протекает бессимптомно, но острая тяжелая легочная инфекция возникает либо у иммуносупрессивного хозяина, либо у иммунокомпетентного хозяина, который вдыхает большой инокулят клеток H. capsulatum . Произошли вспышки от нескольких до десятков тысяч человек (Бродский и др., 1973; Пшеница и др., 1981; Пшеница, 1997).
Гифальная форма H. capsulatum подвергается бесполому спороношению, в результате чего возникают по крайней мере два типа конидий, макро- и микроконидии (), которые различаются в основном по размеру (Pine 1960).Размер микроконидий составляет от 2 до 6 мкм, тогда как размер макроконидий, как сообщается, составляет от 8 до 14 мкм до 10–25 мкм, в зависимости от штамма и условий роста. Считается, что микроконидии имеют подходящий размер для размещения в альвеолах легких и, таким образом, представляют собой наиболее распространенный заразный рост. Конидии и / или фрагменты гиф вдыхаются хозяином, а затем поглощаются макрофагами и другими фагоцитарными клетками (Eissenberg and Goldman 1991; Bullock 1993; Woods 2003).Попадая внутрь хозяина, и споры, и волокна дают начало дрожжевым клеткам, которые избегают фагоцитарного уничтожения и размножаются в альвеолярных макрофагах. Впоследствии дрожжевые клетки используют фагоцитарные клетки в качестве транспортных средств для распространения во многие органы ретикулоэндотелиальной системы, такие как селезенка, печень, лимфатические узлы и костный мозг. Считается, что гематогенное распространение из легких через инфицированные макрофаги происходит даже при бессимптомных инфекциях (Kauffman 2007). У пациентов с диссеминированным заболеванием могут быть колонизированы другие органы, такие как кожа, сердце, мозг, надпочечники и желудочно-кишечный тракт.У большинства хозяев клеточный иммунитет и соответствующая активация макрофагов служат для сдерживания инфекции (Eissenberg and Goldman 1991; Newman 1999; Deepe 2000; Huffnagle and Deepe 2003). Тем не менее, H. capsulatum может оставаться латентным и реактивироваться спустя годы после первоначального заражения (Kauffman 2007). Пациенты с особым риском диссеминированного гистоплазмоза включают лиц с недостаточным клеточным иммунитетом, включая младенцев, пациентов со СПИДом, реципиентов трансплантатов, пациентов с гематологическими злокачественными новообразованиями и пациентов, проходящих лечение кортикостероидами или антагонистами фактора некроза опухолей (Kauffman 2007; Smith and Kauffman 2009 г.).
Трансформация клеток гиф H. capsulatum в дрожжевые клетки или наоборот может быть воспроизведена в культуре, просто изменяя температуру роста (Maresca and Kobayashi 1989; Maresca et al. 1994). Когда клетки H. capsulatum выращивают при комнатной температуре, они растут в форме гиф. Когда эти клетки перемещаются до 37 ° C, они переходят в форму почкующихся дрожжей. Ряд биологических анализов был применен к дрожжевым и гифальным клеткам, выращенным в лаборатории.Первые факторов вирулентности H. capsulatum (Cbp1 [кальций-связывающий белок 1] и Yps3 [ген 3, специфичный для дрожжевой фазы]) были идентифицированы путем поиска высокопроизводительных секретируемых факторов, которые продуцируются только дрожжевыми клетками (Keath and Abidi 1994; Batanghari and Goldman 1997; Batanghari et al. 1998; Patel et al. 1998; Kugler et al. 2000a, b; Sebghati et al. 2000; Bohse and Woods 2007). Основываясь на надежной системе внехромосомных плазмид (Woods and Goldman 1992, 1993) и технологиях электропорации (Woods et al.1998a), сочетание разрушения гена (которое неэффективно в H. capsulatum из-за, по крайней мере частично, высокой частоты незаконной рекомбинации) (Woods et al. 1998b; Sebghati et al. 2000), Agrobacterium -опосредованный инсерционный мутагенез (Sullivan et al. 2002; Youseff et al. 2009) и РНК-интерференция (Rappleye et al. 2004) были использованы для идентификации факторов H. capsulatum , участвующих в приобретении железа и гомеостазе (Hilty et al. 2008). , 2011; Hwang et al.2008, 2012), внутриклеточный паразитизм (Sebghati et al. 2000; Edwards et al. 2011b; Isaac et al. 2013), синтез α-глюкана клеточной стенки (Rappleye et al. 2004; Marion et al. 2006; Edwards et al. 2011a), приобретение витаминов (Garfoot et al. 2014) и детоксикация супероксидом (Youseff et al. 2012; Holbrook et al. 2013). В дополнение к этим молекулярно-генетическим подходам был использован ряд подходов геномики и протеомики для определения генов, экспрессия которых повышена в дрожжах или гифах (Hwang et al.2003; Нгуен и Сил 2008; Beyhan et al. 2013; Эдвардс и др. 2013) и H. capsulatum, секретируемый протеом (Albuquerque et al. 2008; Holbrook et al. 2011), который предположительно включает новые факторы вирулентности. Наконец, были идентифицированы ключевые регуляторные молекулы, необходимые для зависимого от температуры переключения от гифы к дрожжевым формам (Nemecek et al. 2006; Nguyen and Sil 2008; Webster and Sil 2008; Beyhan et al. 2013). Эти факторы прольют свет на механизм определения температуры, который является ключом к диморфному переключению у этих организмов.
Paracoccidioides brasiliensis
P. brasiliensis является возбудителем паракокцидиоидомикоза, заболевания, впервые описанного в 1908 году (см. Goldani and Sugar 1995). Это наиболее распространенный системный микоз в Латинской Америке, число инфицированных составляет около 10 миллионов человек (Bethlem et al. 1991; Brummer et al. 1993). P. brasiliensis как вид, как было показано, состоит из ряда отдельных филогенетических линий, которые недавно были разделены на два вида: P.brasiliensis и P. lutzii (Matute et al. 2006a, b). Paracoccidioides считается первичным патогеном, поражающим людей, считающихся в остальном здоровыми. Однако инфекции чаще всего протекают бессимптомно, и считается, что во многих из этих случаев грибок остается в спящем состоянии. Прогрессирование заболевания, вероятно, связано с некоторой формой иммунодефицита у хозяина (Franco et al. 1987). Озадачивающим аспектом паракокцидиоидомикоза является очевидная низкая распространенность СПИДа, несмотря на высокую частоту сосуществования с ВИЧ-инфекцией в некоторых регионах.Частично это объяснение может относиться к обычному профилактическому применению триметприм-сульфаметоксазола против Pneumocystis carinii ; этот препарат также эффективен против P. brasiliensis . Другая возможность, характерная для многих грибковых инфекций, — это плохое выявление или неправильный диагноз в сочетании с тем фактом, что вскрытие трупов, погибших от СПИДа, обычно не проводится (Goldani and Sugar 1995). Паракокцидиоидомикоз обычно описывается как хроническое гранулематозное воспаление, но у отдельных пациентов может проявляться ряд клинических проявлений, начиная от локализованного доброкачественного заболевания до диссеминированного системного микоза.Распространенное заболевание часто приводит к летальному исходу, если его не лечить (Franco et al. 1987).
P. brasiliensis является эндемиком и встречается только в Южной и Центральной Америке. Основным резервуаром окружающей среды, идентифицированным на сегодняшний день, являются эндемичные для этого региона броненосцы, но они также были обнаружены у ряда других млекопитающих (Richini-Pereira et al. 2008). Высокая ассоциация между Paracoccidioides и броненосцами ( Dasypus septemcinctus ), температура тела которых составляет 32–35 ° C, имеют предположительно слабую иммунную систему и являются роющими животными, что может лежать в основе эволюции патогенности у Paracoccidioides .Грибок также был спорадически изолирован из ряда других источников, включая гуано летучих мышей и пингвинов (обзор в Bagagli et al. 2008). Тесты роста в различных безвредных условиях показывают явное предпочтение определенных почв с высоким содержанием воды.
Диморфный переключатель в P. brasiliensis также срабатывает по температуре. При 25 ° C образуются удлиненные клетки гиф, которые растут апикально и делятся септацией. Субапикальные ветви образуют мицелиальную сеть. Внутри этой сети клеток гиф бесполые конидии образуются без развития сложной структуры конидиофор (см. Goldani and Sugar 1995).Кроме того, также наблюдается артроконидиация () (Bagagli et al. 2008). При 37 ° C дрожжевые клетки делятся почкованием, но полное разделение материнских и дочерних клеток происходит медленно. Это приводит к типичному расположению клеток с множеством бутонов (Goldani and Sugar 1995). Было высказано предположение, что это большое скопление клеток невосприимчиво к фагоцитозу, и это согласуется с наблюдением, что клетки P. brasiliensis в организме хозяина обычно не являются внутриклеточными. Однако также ясно, что вдыхаемые конидии, вероятно, будут фагоцитированы альвеолярными макрофагами, прежде чем они смогут прорасти и продуцировать эти многопучковые формы дрожжей.Переход от гифы к дрожжевым клеткам in vitro происходит медленно, для полного проявления требуется 10–20 дней. Хотя телеоморфная стадия не описана, имеются убедительные доказательства полового цикла у некоторых изолятов (основанные на рекомбинации), тогда как другие кажутся бесполыми (Silva et al. 2008).
Paracoccidioides Предполагается, что инфекция возникает при вдыхании конидий или артроконидий и в основном происходит в легких. Были отмечены некоторые наблюдения Paracoccidioides внутри клеток-хозяев (эпителиальных и альвеолярных), и считается, что первоначальное выживание гриба в макрофагах может быть важным для вирулентности (Tavares et al.2007). Тем не менее, Paracoccidioides не является преимущественно внутриклеточным патогеном; предположительно, как упоминалось выше, форма многопучковых дрожжей слишком велика, чтобы легко фагоцитироваться клетками врожденного иммунитета (Mendes-Giannini et al. 2008). Как внеклеточный патоген, важно прикрепление к клеткам-хозяевам. Инфекции могут быть бессимптомными или симптоматическими, и в первом случае болезнь может проявиться через много лет после первоначального заражения, что указывает на латентный период. Помимо первичного легочного заболевания, диссеминированное заболевание также может проявляться, поражая ретикулоэндотелиальную систему, лимфатические узлы, кожу и слизистую оболочку (Goldani and Sugar 1995).
Разработаны молекулярно-генетические инструменты для обработки P. brasiliensis . Система ДНК-опосредованной трансформации основана на системе трансформации Т-ДНК Agrobacterium , разработанной для H. capsulatum с использованием доминантного селектируемого маркера устойчивости к гигромицину (Almeida et al. 2007). Система была соединена с конструкциями антисмысловой РНК для подавления экспрессии генов. Нацеливание на гены еще не разработано. Было показано, что гены флуоресцентных белков работают у P.brasiliensis (Алмейда и др., 2009). Существуют надежные клеточные и мышиные модели для оценки вирулентности (Defaveri et al. 1982; Kerr et al. 1982; Robledo et al. 1982). Транскрипционное профилирование с использованием микроматриц широко используется (Nunes et al. 2005; Andrade et al. 2006; Ferreira et al. 2006; Tavares et al. 2007; Monteiro et al. 2009).
Sporothrix schenckii
S. schenckii — гриб, распространенный по всему миру, в отличие от многих других диморфных патогенов и больше похожий на более известный, но недиморфный гриб Aspergillus fumigatus.S. schenckii ассоциируется с почвой и растениями. В отличие от других диморфных патогенов, S. schenckii преимущественно вызывает кожные инфекции, которые инициируются незначительными травмами кожи, такими как порезы и царапины, возникающие при работе с инфицированным материалом. Это заболевание часто называют болезнью цветоводов. Впервые о споротрихозе сообщили в 1896 году, когда грибок был обнаружен в подкожных абсцессах (Schenck 1898). Особенно печально известная вспышка споротрихоза произошла в Южной Африке, где 3000 золотодобытчиков были заражены зараженной древесиной (цитируется по Lopez-Romero et al.2011). Заболевание часто проявляется в виде хронической гранулематозной инфекции. Как и Paracoccidioides , S. schenckii теперь считается комплексом видов, состоящим из шести видов, каждый из которых представляет медицинский интерес, и, если не указано иное, ссылка на S. schenckii относится к комплексу видов. (Лопес-Ромеро и др., 2011).
Несмотря на глобальное распространение, некоторые регионы, такие как Перу, по-видимому, демонстрируют гораздо более высокие уровни инфицирования на S.schenckii (иногда называемый «гиперэндемическим»), вероятно, как следствие более высокой частоты активности, способствующей инфицированию (Pappas et al. 2000). Считается, что контакт также является причиной гораздо более высокой частоты инфицирования мужчин S. schenckii . Он присутствует в разлагающемся (или нет) растительном материале. Ряд факторов способствует развитию споротрихоза, включая размер и расположение инокулята, иммунное состояние хозяина и вирулентность штамма (Dixon et al.1991). Обычно инфекции ограничиваются острыми или хроническими подкожными поражениями, но серьезное диссеминированное заболевание очевидно с увеличением числа лиц с ослабленным иммунитетом, особенно СПИДом (Lopez-Romero et al. 2011). Также задокументировано заражение от животных, таких как кошки (Barros et al. 2004).
Sporothrix отличается от других диморфных грибов тем, что клетки гиф и дрожжевые клетки могут сосуществовать, а температура, по-видимому, не является строгим признаком перехода между двумя типами клеток.Клетки гифы растут апикальным расширением, делятся перегородкой и образуют ряд морфологически различных бесполых конидий. Некоторые конидии имеют овальную или удлиненную форму, которые могут быть получены индивидуально или небольшими группами симпоидным способом из специализированных конидиогенных клеток. Другой тип конидиальных клеток имеет толстую стенку, темную пигментацию и, как правило, продуцируется индивидуально. Этот тип конидий отличается по форме от видов Sporothrix . Sporothrix дрожжевые клетки имеют овальную форму и делятся почкованием.Разделение клеток может происходить при каждом делении или может быть отложено для получения многослойных клеток. Эти клетки могут продуцироваться в диапазоне температур (25–37 ° C) и обычно появляются из клеток гиф на их апикальных концах или в местах перегородок. Половая стадия неизвестна (Lopez-Romero et al. 2011).
Для S. schenckii , в отличие от всех других диморфных патогенов, обсуждаемых здесь, основным путем заражения является не вдыхание спор (хотя есть доказательства такой возможности), а вместо этого через гифальную форму при инокуляции через поверхностное ранение.Инфекции обычно локализуются на коже, в результате чего образуются изъязвленные узелки, а иногда распространяются на лимфатическую систему. У людей с ослабленным иммунитетом, например, больных СПИДом, часто встречается серьезное диссеминированное заболевание, которое может поражать многие части тела, включая кости, суставы и центральную нервную систему (Lopez-Romero et al. 2011).
Для Sporothrix имеется мало молекулярно-генетических инструментов. Система ДНК-опосредованной трансформации с использованием инсерции, опосредованной Т-ДНК Agrobacterium , была разработана на основе системы, описанной для T.marneffei и Aspergillus awamori и использовали для скрининга инсерционного мутагенеза (Zhang et al. 2011). Нацеливание на гены еще не разработано. Гель-электрофорез в импульсном поле использовался для изучения хромосомного полиморфизма изолятов (Sasaki et al. 2014). Существуют надежные клеточные и мышиные модели для оценки вирулентности (Hachisuka and Sasai 1981; Kennedy et al. 1982; Dickerson et al. 1983).
Talaromyces marneffei
Penicillium marneffei , недавно переименованный в T.marneffei на основе новых данных о последовательностях, которые видели разделение классически определенной группы грибов Penicillium (Samson et al. 2011), был первоначально идентифицирован в 1956 году от бамбуковой крысы во Вьетнаме (Segretain 1959). Несмотря на его сосуществование с крысами в эндемичных регионах, его редко изолировали от окружающей среды, что позволяет предположить, что резервуаром на самом деле может быть крыса. Первое задокументированное заражение человека произошло в результате случайного укола иглой исследователя (DiSalvo et al. 1973).Он приобрел известность как значительный патоген для человека в Юго-Восточной Азии в связи с глобальной пандемией СПИДа (Vanittanakom et al. 2006). Заболевание penicilliosis marneffei возникает преимущественно у лиц с ослабленным иммунитетом и начинается как легочная инфекция с последующим гематогенным распространением в системный микоз (Cheng et al. 1998; Rimek et al. 1999; Garbino et al. 2001). Таким образом, он считается условно-патогенным микроорганизмом; однако системное заболевание без лечения фатально. Было описано небольшое количество случаев инфицирования «иммунокомпетентных» людей, но следует отметить, что ни в одном из этих случаев иммунный статус не был должным образом протестирован, а «иммунокомпетентность» часто используется как взаимозаменяемые (и неправильно) с ВИЧ-отрицательными. статус.Британскому писателю и писателю-путешественнику Брюсу Чатвину (1963–1989) был поставлен диагноз «марнефейный пенициллиоз» (Shakespeare and Neely 2011).
T. marneffei эндемичен для Юго-Восточной Азии и вследствие растущей заболеваемости представляет собой «СПИД-определяющий патоген» в этом регионе (Supparatpinyo et al. 1994; Ustianowski et al. 2008). Увеличивающаяся частота поездок в Азию и способность T. marneffei существовать без хозяина привели к распространению инцидентов по Азии и подтвержденным случаям в Европе, Северной Америке, Африке и Австралии (Vanittanakom et al.2006 г.). Несмотря на свою прототрофность и способность расти на широком диапазоне источников питательных веществ, что свидетельствует о сапрофитном образе жизни в форме гиф, T. marneffei почти всегда ассоциируется с бамбуковыми крысами в окружающей среде (см. Обзор в Vanittanakom et al. 2006 ). Частично причина этого может заключаться в том, что он относительно медленно растет и может плохо конкурировать с другими микробами в почве (Vanittanakom et al. 1995; Joshi et al. 2003). Бамбуковая крыса как экологическая ниша, возможно, способствовала развитию ее патогенного потенциала, и неясно, обнаруживается ли форма роста дрожжей вне млекопитающих-хозяев.Популяционно-генетические исследования изолятов от бамбуковых крыс и людей в Азии предполагают высококлональную структуру, но с явными признаками рекомбинации. Эти популяции кажутся пространственно ограниченными и пересекаются с характерным распределением бамбуковых крыс. В сочетании с данными экспериментов на мышах оказывается, что рекомбинация может происходить внутри хозяев, и, если это необходимо, она может представлять популяционные барьеры для T. marneffei (Henk et al. 2012). Нет никаких свидетельств того, что T.marneffei инфекции у людей происходят от зоонозов, и общепринято считать, что, как и у многих других грибов, инфекция инициируется вдыханием спящих конидий. Это подтверждают данные, связывающие инфекции с занятием сельским хозяйством и увеличением заболеваемости в сезон дождей, по крайней мере, в Северном Таиланде (Chariyalertsak et al. 1996).
Как и другие важные диморфные грибковые патогены, T. marneffei демонстрирует диморфный рост с образованием двух различных клеточных форм, одноклеточных дрожжей и многоклеточных гиф, в определенных условиях окружающей среды (Andrianopoulos 2002). T. marneffei — единственный известный вид Talaromyces ( Penicillium ), который является диморфным или патогеном человека, что позволяет предположить, что они связаны между собой возникающими признаками. Переключение между формами роста регулируется температурой (Андрианопулос, 2002). При 25 ° C T. marneffei растет в виде многоядерных, перегородчатых, разветвленных гиф. Эти гифы продуцируют конидии, инфекционный агент, из специализированных многоклеточных структур, называемых конидиеносными. При переключении на 37 ° C Т.marneffei претерпевает морфогенетический процесс, называемый артроконидиацией. Клеточное и ядерное деление становятся связанными, закладываются двойные перегородки, и гифы фрагментируются в этих местах перегородки, чтобы высвободить одноядерные дрожжевые клетки, которые делятся путем деления (Andrianopoulos 2002). Клетки дрожжей являются патогенной формой и обнаруживаются в легочных альвеолярных макрофагах и мононуклеарных клетках периферической крови инфицированных людей (Vanittanakom et al. 2006). В отличие от диморфного перехода гифы и дрожжей in vitro, конидии, фагоцитируемые макрофагами, прорастают непосредственно в дрожжевые клетки, минуя процесс артроконидиации.Внутриклеточные дрожжевые клетки морфологически сходны с таковыми других внутриклеточных грибковых патогенов, таких как H. capsulatum , но их можно различить при делении, поскольку дрожжевых клеток T. marneffei делятся путем деления (Игнатов и Кит, 2002).
Инфекция T. marneffei , вероятно, происходит при вдыхании конидий, которые фагоцитируются легочными альвеолярными макрофагами хозяина (Vanittanakom et al. 2006). Как T. marneffei может выжить в стрессовой внутриклеточной среде фагоцитарного макрофага, не совсем ясно.Конидии, фагоцитируемые альвеолярными макрофагами, прорастают внутри этих клеток-хозяев в патогенную дрожжевую форму и размножаются. Это приводит к легочной инфекции, которая гемотогенно распространяется в лимфатическую систему, печень, селезенку и кости (Kudeken et al. 1996). Поражения кожи часто проявляются в основном на лице и шее. Нелеченная диссеминированная болезнь часто заканчивается летальным исходом. Эксперименты на мышах показали, что в отсутствие иммуносупрессии инфекции с большим количеством конидий (4 × 10 5 ) в конечном итоге исчезают, тогда как бестимусные мыши умирают от инфекции (Kudeken et al.1997). Это показывает, что иммунный статус хозяина является ключевым фактором исхода инфекции.
Обширный набор молекулярно-генетических инструментов был разработан для T. marneffei с использованием типового штамма (FRR2161 / ATCC18224). Эти инструменты использовались для исследования молекулярных механизмов, которые контролируют диморфный переключатель и позволяют грибку выжить в организме хозяина. К ним относятся (1) очень высокочастотная процедура ДНК-опосредованной трансформации с использованием слияния протопластов на основе ПЭГ, которая приводит к интеграции экзогенной ДНК (Borneman et al.2001), (2) набор питательных и доминантных селектируемых маркеров с соответствующими штаммами-реципиентами для трансформации (Boyce et al. 2012), (3) усиленные штаммы, в которых интеграция ДНК осуществляется строго путем гомологичной рекомбинации с использованием мутантов в негомологичных концевых соединениях. (NHEJ) система (Bugeja et al. 2012), (4) штаммы, экспрессирующие флуоресцентные белки для отслеживания живых клеток (Boyce et al. 2012), (5) системы нокдауна антисмысловой РНК (Canovas et al. 2011), (6) системы шлюзов для быстрого создания конструкций (Boyce et al.2012; Bugeja et al. 2012), (7) промоторы для контролируемой экспрессии генов (Borneman et al. 2000; Boyce et al. 2001), (8) геномные и транскриптомные технологии (Pasricha et al. 2013) и (9) культура клеток рыбок данио, и мышиные системы для тестирования вирулентности (Boyce and Andrianopoulos 2007; Ellett et al. 2011; Henk et al. 2012). Также описана процедура ДНК-опосредованной трансформации с использованием системы передачи ДНК Agrobacterium (Kummasook et al. 2010).
ЗАКЛЮЧЕНИЕ
Термически диморфные грибы представляют собой увлекательную группу организмов, которые используют множество стратегий для управления прогрессированием болезни у хозяина.Эволюционная связь между диморфизмом и вирулентностью является предметом большого интереса для микробиологов, а появление надежных молекулярных подходов и секвенирования следующего поколения делают это благоприятным временем для изучения патогенеза и базовой биологии этих удивительных организмов.
Сноски
Редакторы: Артуро Касадеваль, Аарон П. Митчелл, Джудит Берман, Кьюнг Дж. Квон-Чунг, Джон Р. Перфект и Джозеф Хейтман
Дополнительные перспективы по патогенам человеческих грибков доступны на сайте www.Перспективыinmedicine.org
ССЫЛКИ
- Abuodeh RO, Orbach MJ, Mandel MA, Das A, Galgiani JN. 2000 г.
Генетическая трансформация Coccidioides immitis при содействии Agrobacterium tumefaciens . J заразить Дис
181: 2106–2110. [PubMed] [Google Scholar] - Albuquerque PC, Nakayasu ES, Rodrigues ML, Frases S, Casadevall A, Zancope-Oliveira RM, Almeida IC, Nosanchuk JD. 2008 г.
Везикулярный транспорт в Histoplasma capsulatum : эффективный механизм переноса белков и липидов trans через стенку клетки у аскомицетов.Клеточный микробиол
10: 1695–1710. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Almeida AJ, Carmona JA, Cunha C, Carvalho A, Rappleye CA, Goldman WE, Hooykaas PJ, Leao C, Ludovico P, Rodrigues F. 2007.
На пути к молекулярно-генетической системе патогенного гриба Paracoccidioides brasiliensis . Грибок Genet Biol
44: 1387–1398. [PubMed] [Google Scholar] - Алмейда А.Дж., Кунья С., Кармона Д.А., Сампайо-Маркес Б., Карвалью А., Малавази И., Стинсма Х.Й., Джонсон Д.И., Леао С., Логаринхо Э. и др.2009 г.
Cdc42p контролирует форму дрожжевых клеток и вирулентность Paracoccidioides brasiliensis . Грибок Genet Biol
46: 919–926. [PubMed] [Google Scholar] - Андраде Р.В., Паес ХК, Никола А.М., де Карвалью М.Дж., Фачин А.Л., Кардосо Р.С., Сильва С.С., Фернандес Л., Силва С.П., Донади Е.А. и др.
2006 г.
Гены, связанные с организацией клеток, метаболизмом серы и переносом ионов, по-разному экспрессируются в мицелии Paracoccidioides brasiliensis и дрожжевых клетках. BMC Genomics
7: 208.[Бесплатная статья PMC] [PubMed] [Google Scholar] - Andrianopoulos A. 2002.
Контроль морфогенеза грибкового патогена человека Penicillium marneffei . Int J Med Microbiol
292: 331–347. [PubMed] [Google Scholar] - Арнетт М.В., Фрейзер С.Л., Грбах В.Х. 2008 г.
На Гавайях диагностирован легочный бластомикоз. Юго-Восточная Азия J Trop Med Public Health
39: 701–705. [PubMed] [Google Scholar] - Ashburn LL, Emmons CW. 1942 г.
Спонтанная кокцидиоидная гранулема в легких диких грызунов.Арка Патол
34: 791–800. [Google Scholar] - Bagagli E, Theodoro RC, Bosco SM, McEwen JG. 2008 г.
Paracoccidioides brasiliensis : Филогенетические и экологические аспекты. Микопатология
165: 197–207. [PubMed] [Google Scholar] - Баррос М.Б., Шубах Аде О, до Валле А.С., Гутьеррес Гальхардо М.С., Консейсао-Сильва Ф., Шубах Т.М., Рейс Р.С., Ванке Б., Марзочи К.Б., Консейсао М.Дж. 2004 г.
Эпидемия споротрихоза, передаваемого кошками, в Рио-де-Жанейро, Бразилия: описание серии случаев.Clin Infect Dis
38: 529–535. [PubMed] [Google Scholar] - Батангхари JW, Goldman WE. 1997 г.
Кальциевая зависимость и связывание в культурах Histoplasma capsulatum . Заразить иммунную
65: 5257–5261. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Batanghari JW, Deepe GS Jr, Di Cera E, Goldman WE. 1998 г.
Получение гистоплазмой кальция и экспрессия CBP1 во время внутриклеточного паразитизма. Мол Микробиол
27: 531–539. [PubMed] [Google Scholar] - Баумгарднер Д.П., Парецкий Д.П., Йопп А.С.1995 г.
Эпидемиология бластомикоза у собак: Северо-центральный Висконсин, США. J Med Vet Mycol
33: 171–176. [PubMed] [Google Scholar] - Bethlem NM, Lemle A, Bethlem E, Wanke B. 1991.
Паракокцидиоидомикоз. Семин Респир Мед
12: 81–86. [Google Scholar] - Бейхан С., Гутьеррес М., Вурхис М., Сил А. 2013.
Сеть, чувствительная к температуре, связывает форму клеток и признаки вирулентности первичного грибкового патогена. PLoS Biol
11: e1001614. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Bohse ML, Woods JP.2007 г.
Опосредованное РНК-интерференцией молчание гена YPS3 из Histoplasma capsulatum выявляет дефекты вирулентности. Заразить иммунную
75: 2811–2817. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Borneman AR, Hynes MJ, Andrianopoulos A. 2000.
Гомолог abaA из Penicillium marneffei участвует в двух программах развития: конидиации и диморфном росте. Мол Микробиол
38: 1034–1047. [PubMed] [Google Scholar] - Борнеман А.Р., Хайнс М.Дж., Андрианопулос А.2001 г.
Гомолог STE12 из бесполого диморфного гриба Penicillium marneffei дополняет дефект полового развития мутанта Aspergillus nidulans steA . Генетика
157: 1003–1014. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Boyce KJ, Andrianopoulos A. 2007.
Активированная p21 киназа необходима для прорастания конидий в Penicillium marneffei . PLoS Pathog
3: e162. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Boyce KJ, Hynes MJ, Andrianopoulos A.2001 г.
Гомолог CDC42 диморфного гриба Penicillium marneffei необходим для правильной поляризации клеток во время роста, но не развития. J Бактериол
183: 3447–3457. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Boyce KJ, Bugeja HE, Weerasinghe H, Payne M, Schreider L, Park C, Woodward T, Andrianopoulos A. 2012.
Стратегии молекулярно-генетической манипуляции и визуализации грибкового патогена человека Penicillium marneffei . Грибной Genet Rep
59: 1–12.[Google Scholar] - Brandhorst TT, Rooney PJ, Sullivan TD, Klein BS. 2002 г.
Использование новых генетических инструментов для изучения патогенеза Blastomyces dermatitidis . Тенденции Microbiol
10: 25–30. [PubMed] [Google Scholar] - Бродский А.Л., Грегг М.Б., Лёвенштейн М.С., Кауфман Л., Маллисон Г.Ф. 1973 г.
Вспышка гистоплазмоза, связанная с мероприятиями в честь Дня Земли 1970 года. Am J Med
54: 333–342. [PubMed] [Google Scholar] - Brown GD, Denning DW, Gow NA, Levitz SM, Netea MG, White TC.2012 г.
Скрытые убийцы: грибковые инфекции человека. Sci Transl Med
4: 165rv113. [PubMed] [Google Scholar] - Браммер Э., Кастанеда Э., Рестрепо А. 1993.
Паракокцидиоидомикоз: обновление. Clin Microbiol Rev
6: 89–117. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Bugeja HE, Boyce KJ, Weerasinghe H, Beard S, Jeziorowski A, Pasricha S, Payne M, Schreider L, Andrianopoulos A. 2012.
Инструменты для высокоэффективной генетической манипуляции с патогеном человека Penicillium marneffei .Грибок Genet Biol
49: 772–778. [PubMed] [Google Scholar] - Bullock WE. 1993 г.
Взаимодействие между фагоцитарными клетками человека и Histoplasma capsulatum . Arch Med Res
24: 219–223. [PubMed] [Google Scholar] - Кано М.В., Понсе-де-Леон Г.Ф., Типпен С., Линдсли М.Д., Уорвик М., Хаджех Р.А. 2003 г.
Бластомикоз в Миссури: эпидемиология и факторы риска эндемических заболеваний. Эпидемиол Инфекция
131: 907–914. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Canovas D, Boyce KJ, Andrianopoulos A.2011 г.
Грибковый миозин типа II в Penicillium marneffei , MyoB, важен для отложения хитина в участках зарождающейся перегородки, но не для локализации актина. Эукариотическая клетка
10: 302–312. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Chariyalertsak S, Sirisanthana T, Supparatpinyo K, Nelson KE. 1996 г.
Сезонная изменчивость распространенных инфекций Penicillium marneffei в северном Таиланде: ключ к резервуару?
J заразить Дис
173: 1490–1493. [PubMed] [Google Scholar] - Cheng NC, Wong WW, Fung CP, Liu CY.1998 г.
Необычные легочные проявления диссеминированной инфекции Penicillium marneffei у трех больных СПИДом. Мед Микол
36: 429–432. [PubMed] [Google Scholar] - Chiller TM, Галгиани Дж. Н., Стивенс Д. А.. 2003 г.
Кокцидиоидомикоз. Инфекция Dis Clin North Am
17: 41–57. [PubMed] [Google Scholar] - Чу Дж. Х., Фейдтнер К., Хейдон К., Уолш Т. Дж., Заутис Т. Е.. 2006 г.
Госпитализации по поводу эндемических микозов: национальное популяционное исследование. Clin Infect Dis
42: 822–825. [PubMed] [Google Scholar] - Кокшотт В.П., Лукас А.О.1964 г.
Гистоплазмоз дубоизии. Q J Med
33: 223–238. [PubMed] [Google Scholar] - Converse JL. 1955 г.
Рост шариков Coccidioides immitis в химически определенной жидкой среде. Proc Soc Exp Biol Med
90: 709–711. [PubMed] [Google Scholar] - Crampton TL, Light RB, Berg GM, Meyers MP, Schroeder GC, Hershfield ES, Embil JM. 2002 г.
Эпидемиология и клинический спектр бластомикоза, диагностированного в больницах Манитобы. Clin Infect Dis
34: 1310–1316. [PubMed] [Google Scholar] - Darling ST.1906 г.
Общая протозойная инфекция, вызывающая псевдотуберкулы в легких и очаговые некрозы в печени, селезенке и лимфатических узлах. JAMA
46: 1283–1285. [Google Scholar] - Deepe GS Jr. 2000.
Иммунный ответ на ранние и поздние инфекции Histoplasma capsulatum . Curr Opin Microbiol
3: 359–362. [PubMed] [Google Scholar] - Дефавери Дж., Резкаллах-Ивассо М.Т., де Франко М.Ф. 1982 г.
Экспериментальный легочный паракокцидиоидомикоз у мышей: морфология и корреляция поражений с гуморальным и клеточным иммунным ответом.Микопатология
77: 3–11. [PubMed] [Google Scholar] - Де Гроот М.А., Бьерке Р., Смит Х., Родос, Иллинойс. 2000 г.
Расширение эпидемиологии бластомикоза: клинические особенности и расследование 2 случаев в Колорадо. Clin Infect Dis
30: 582–584. [PubMed] [Google Scholar] - Дикерсон К.Л., Тейлор Р.Л., Drutz DJ. 1983 г.
Восприимчивость врожденно бестимусных (голых) мышей к споротрихозу. Заразить иммунную
40: 417–420. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Dickson EC. 1915 г.
Оидомикоз в Калифорнии с особым упором на кокцидиоидную гранулему, включая девять новых случаев кокцидиоидной гранулемы.Arch Internal Med
15: 479–486. [Google Scholar] - DiSalvo AF, Fickling AM, Ajello L. 1973.
Инфекция, вызванная Penicillium marneffei : Описание первой естественной инфекции у человека. Am J Clin Pathol
60: 259–263. [PubMed] [Google Scholar] - Диксон Д.М., Салкин И.Ф., Дункан Р.А., Херд, штат Нью-Джерси, Хейнс Дж. Х., Кемна М. Е., Коулс Ф. Б. 1991 г.
Выделение и характеристика Sporothrix schenckii из клинических и экологических источников, связанных с крупнейшей эпидемией споротрихоза в США.J Clin Microbiol
29: 1106–1113. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Dworkin MS, Duckro AN, Proia L, Semel JD, Huhn G. 2005.
Эпидемиология бластомикоза в Иллинойсе и факторы, связанные со смертью. Clin Infect Dis
41: e107 – e111. [PubMed] [Google Scholar] - Эдвардс Дж. А., Алоре Е. А., Рэппли, Калифорния. 2011a.
Требования к вирулентности дрожжевой фазы для α-глюкансинтазы различаются для хемотипов Histoplasma capsulatum . Эукариотическая клетка
10: 87–97. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Edwards JA, Zemska O, Rappleye CA.2011b.
Обнаружение роли Hsp82 в вирулентности Histoplasma посредством количественного скрининга летальности макрофагов. Заразить иммунную
79: 3348–3357. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Эдвардс JA, Chen C, Kemski MM, Hu J, Mitchell TK, Rappleye CA. 2013.
Histoplasma дрожжевые и мицелиальные транскриптомы выявляют профили экспрессии генов, специфичных для патогенной фазы и клон-специфического происхождения. BMC Genomics
14: 695. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Eissenberg LG, Goldman WE.1991 г.
Histoplasma Вариации и адаптивные стратегии для паразитизма: новые взгляды на гистоплазмоз. Clin Microbiol Rev
4: 411–421. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Эллетт Ф., Пазе Л., Хейман Дж. У., Андрианопулос А., Лишке Дж. Дж. 2011 г.
mpeg1 Промоторные трансгены направляют экспрессию клонов макрофагов у рыбок данио. Кровь
117: e49 – e56. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Emmons CW. 1949 г.
Выделение Histoplasma capsulatum из почвы.Представитель общественного здравоохранения
64: 892–896. [PubMed] [Google Scholar] - Эммонс CW. 1962 г.
Почвенные резервуары патогенных грибов. J Wash Acad Sci
52: 3–9. [Google Scholar] - Феррейра М.Э., Маркес Эдос Р., Малавази И., Торрес И., Рестрепо А., Нуньес Л. Р., де Оливейра Р. К., Голдман М. Х., Голдман Г. Х. 2006 г.
Транскриптомный анализ и молекулярные исследования метаболизма серы в патогенных грибах человека Paracoccidioides brasiliensis . Мол Генет Геномикс
276: 450–463. [PubMed] [Google Scholar] - Фишер М.С., Кениг Г.Л., Уайт Т.Дж., Тейлор Дж.В.2002 г.
Молекулярно-фенотипическое описание Coccidioides posadasii sp. nov., ранее считавшейся некалифорнийской популяцией Coccidioides immitis . Микология
94: 73–84. [PubMed] [Google Scholar] - Franco M, Montenegro MR, Mendes RP, Marques SA, Dillon NL, Mota NG. 1987 г.
Паракокцидиоидомикоз: недавно предложенная классификация его клинических форм. Rev Soc Бюстгальтеры Med Trop
20: 129–132. [PubMed] [Google Scholar] - Гальгиани Дж. Н., Ампель Н. М., Блэр Дж. Э., Катандзаро А., Джонсон Р. Х., Стивенс Д. А., Уильямс П.Л.2005 г.
Кокцидиоидомикоз. Clin Infect Dis
41: 1217–1223. [PubMed] [Google Scholar] - Гарбино Дж., Коларова Л., Лью Д., Хиршель Б., Ронер П. 2001.
Фунгемия у ВИЧ-инфицированных пациентов: 12-летнее исследование в больнице третичного уровня. Лечение больных СПИДом ЗППП
15: 407–410. [PubMed] [Google Scholar] - Garfoot AL, Zemska O, Rappleye CA. 2014 г.
Histoplasma capsulatum зависит от биосинтеза витаминов de novo для внутрифагосомной пролиферации. Заразить иммунную
82: 393–404. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Готье GM, Салливан Т.Д., Галлардо С.С., Брандхорст Т.Т., Ванден Вимеленберг А.Дж., Куомо Калифорния, Суен Джи, Карри К.Р., Кляйн Б.С.2010 г.
SREB, фактор транскрипции GATA, который управляет разными судьбами в Blastomyces dermatitidis , включая морфогенез и биосинтез сидерофоров. PLoS Pathog
6: e1000846. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Gilchrist TC. 1894 г.
Protazoan dermatitidis. Дж. Кутан Генитурин Дис
12: 496–499. [Google Scholar] - Гилкрист Т.С., Стокс В.Р. 1896 г.
Присутствие Oidium в тканях случая pseudolupus vulgaris. Булл Джонс Хопкинс Больница
7: 129–133.[Google Scholar] - Гилкрист Т.С., Стокс В.Р. 1898 г.
Случай псевдолюпуса, вызванного Blastomyces . J Exp Med
3: 53–78. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Goldani LZ, Sugar AM. 1995 г.
Паракокцидиоидомикоз и СПИД: обзор. Clin Infect Dis
21: 1275–1281. [PubMed] [Google Scholar] - Hachisuka H, Sasai Y.1981.
Развитие экспериментального споротрихоза у нормальных и модифицированных животных. Микопатология
76: 79–82. [PubMed] [Google Scholar] - Hector RF, Rutherford GW, Tsang CA, Erhart LM, McCotter O, Anderson SM, Komatsu K, Tabnak F, Vugia DJ, Yang Y и др.2011 г.
Воздействие кокцидиоидомикоза на общественное здоровье в Аризоне и Калифорнии. Int J Environ Res Public Health
8: 1150–1173. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Хенк Д.А., Шахар-Голан Р., Деви К.Р., Бойс К.Дж., Жан Н., Федорова Н.Д., Нирман В.К., Сюэ П.Р., Юэнь К.Ю., Сиеу Т.П. и др.
2012 г.
Клональность независимо от пола: эволюция связанных с хозяином половых соседей в патогенном грибе Penicillium marneffei . PLoS Pathog
8: e1002851. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hilty J, Smulian AG, Newman SL.2008 г.
Вакуолярная АТФаза Histoplasma capsulatum необходима для гомеостаза железа, внутриклеточной репликации в макрофагах и вирулентности в мышиной модели гистоплазмоза. Мол Микробиол
70: 127–139. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hilty J, George Smulian A, Newman SL. 2011 г.
Histoplasma capsulatum использует сидерофоры для внутриклеточного накопления железа в макрофагах. Мед Микол
49: 633–642. [PubMed] [Google Scholar] - Hoff GL, Bigler WJ.1981 г.
Роль летучих мышей в размножении и распространении гистоплазмоза: обзор. Дж. Уайлдл Дис
17: 191–196. [PubMed] [Google Scholar] - Холбрук Э.Д., Эдвардс Дж. А., Юсефф Б. Х., Рэппли, Калифорния. 2011 г.
Определение внеклеточного протеома патогенной фазы Histoplasma capsulatum . J Proteome Res
10: 1929–1943. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Holbrook ED, Smolnycki KA, Youseff BH, Rappleye CA. 2013.
Избыточные каталазы детоксифицируют реактивный кислород фагоцитов и способствуют патогенезу Histoplasma capsulatum .Заразить иммунную
81: 2334–2346. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Huffnagle GB, Deepe GS. 2003 г.
Врожденные и адаптивные детерминанты восприимчивости хозяев к важным с медицинской точки зрения грибам. Curr Opin Microbiol
6: 344–350. [PubMed] [Google Scholar] - Hung CY, Xue J, Cole GT. 2007 г.
Механизмы вирулентности coccidioides . Энн Нью-Йорк Академия наук
1111: 225–235. [PubMed] [Google Scholar] - Hung CY, Wise HZ, Cole GT. 2012 г.
Нарушение гена в Coccidioides с использованием маркеров устойчивости к гигромицину или флеомицину.Методы Мол Биол
845: 131–147. [PubMed] [Google Scholar] - Хван Л., Хокинг-Мюррей Д., Бахрами А.К., Андерссон М., Райн Дж., Сил А. 2003.
Идентификация фазоспецифических генов грибкового патогена Histoplasma capsulatum с использованием геномного микроматрицы с дробовиком. Mol Biol Cell
14: 2314–2326. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hwang LH, Mayfield JA, Rine J, Sil A. 2008.
Histoplasma требует SID1 , члена кластера генов сидерофоров, регулируемых железом, для колонизации хозяином.PLoS Pathog
4: e1000044. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Hwang LH, Seth E, Gilmore SA, Sil A. 2012.
SRE1 регулирует железозависимые и независимые пути грибкового патогена Histoplasma capsulatum.
Эукариотическая клетка
11: 16–25. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Игнатов А., Кит Э.Дж. 2002 г.
Молекулярная клеточная биология и молекулярная генетика Histoplasma capsulatum . Int J Med Microbiol
292: 349–361. [PubMed] [Google Scholar] - Isaac DT, Coady A, Van Prooyen N, Sil A.2013.
3-гидроксиметилглутарилкофермент А-лиаза HCL1 требуется для колонизации макрофагов грибковым патогеном человека Histoplasma capsulatum . Заразить иммунную
81: 411–420. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Johannesson H, Kasuga T., Schaller RA, Good B, Gardner MJ, Townsend JP, Cole GT, Taylor JW. 2006 г.
Фазоспецифическая экспрессия гена, лежащая в основе морфологической адаптации диморфного патогенного гриба человека, Coccidioides posadasii .Грибок Genet Biol
43: 545–559. [PubMed] [Google Scholar] - Джоши А., Гугнани Х.Г., Виджаян В.К. 2003 г.
Выживание Penicillium marneffei в стерильной и нестерильной почве. J Mycol Med
13: 211–212. [Google Scholar] - Касуга Т., Уайт Т.Дж., Кениг Дж., МакИвен Дж., Рестрепо А., Кастанеда Э., Да Силва Лаказ С., Хейнс-Ваккари Е.М., Де Фрейтас Р.С., Занкоп-Оливейра Р.М. и др.
2003 г.
Филогеография грибкового возбудителя Histoplasma capsulatum . Мол Экол
12: 3383–3401. [PubMed] [Google Scholar] - Kauffman CA.2007 г.
Гистоплазмоз: клинические и лабораторные обновления. Clin Microbiol Rev
20: 115–132. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Keath EJ, Abidi FE. 1994 г.
Молекулярное клонирование и анализ последовательности yps-3 , специфичного для дрожжевой фазы гена диморфного грибкового патогена Histoplasma capsulatum . Микробиология
140: 759–767. [PubMed] [Google Scholar] - Кеннеди MJ, Bajwa PS, Volz PA. 1982 г.
Посев на желудочно-кишечный тракт мышам Sporothrix schenckii .Микопатология
78: 141–143. [PubMed] [Google Scholar] - Kerr IB, da Costa SC, Alencar A. 1982.
Экспериментальный паракокцидиоидомикоз у мышей с подавленным иммунитетом. Immunol Lett
5: 151–154. [PubMed] [Google Scholar] - Киркланд Т.Н., Фирер Дж. 1996.
Кокцидиоидомикоз: возобновляющееся инфекционное заболевание. Emerg Infect Dis
2: 192–199. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Klein BS, Vergeront JM, Davis JP. 1986 г.
Эпидемиологические аспекты бластомикоза, загадочного системного микоза.Семин респир инфекция
1: 29–39. [PubMed] [Google Scholar] - Кляйн Б.С., Верджеронт Дж. М., ДиСальво А. Ф., Кауфман Л., Дэвис Дж. П. 1987 г.
Две вспышки бластомикоза вдоль рек в Висконсине. Выделение Blastomyces dermatitidis из почвы на берегу реки и свидетельства его передачи по водным путям. Am Rev Respir Dis
136: 1333–1338. [PubMed] [Google Scholar] - Krajaejun T, Gauthier GM, Rappleye CA, Sullivan TD, Klein BS. 2007 г.
Разработка и применение системы дозорного зеленого флуоресцентного белка для идентификации РНК-интерференции в Blastomyces dermatitidis проливает свет на роль септина в морфогенезе и споруляции.Эукариотическая клетка
6: 1299–1309. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Кудекен Н., Каваками К., Кусано Н., Сайто А. 1996.
Клеточно-опосредованный иммунитет при устойчивости хозяина к инфекции, вызываемой Penicillium marneffei . J Med Vet Mycol
34: 371–378. [PubMed] [Google Scholar] - Кудекен Н., Каваками К., Сайто А. 1997.
CD4 + Опосредованные Т-клетками фатальные гипервоспалительные реакции у мышей, инфицированных Penicillium marneffei . Клин Эксп Иммунол
107: 468–473.[PubMed] [Google Scholar] - Куглер С., Шурц Себгати Т., Гроппе Айссенберг Л., Голдман В.Е. 2000a.
Фенотипическая изменчивость и внутриклеточный паразитизм по Histoplasma capsulatum . Proc Natl Acad Sci
97: 8794–8798. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Куглер С., Янг Б., Миллер В.Л., Голдман В.Е. 2000b.
Мониторинг фазоспецифической экспрессии гена в Histoplasma capsulatum с теломерными плазмидами слияния GFP. Клеточный микробиол
2: 537–547. [PubMed] [Google Scholar] - Kummasook A, Cooper CR Jr, Ваниттанаком Н.2010 г.
Усовершенствованная система трансформации, опосредованной Agrobacterium, для функционального генетического анализа Penicillium marneffei . Мед Микол
48: 1066–1074. [PubMed] [Google Scholar] - Ларкин М.А., Блэкшилдс Дж., Браун Н.П., Ченна Р., МакГеттиган П.А., МакВильям Х., Валентин Ф., Уоллес И.М., Вильм А., Лопес Р. и др.
2007 г.
Clustal W и Clustal X версии 2.0. Биоинформатика
23: 2947–2948. [PubMed] [Google Scholar] - Lopez-Romero E, Reyes-Montes Mdel R, Perez-Torres A, Ruiz-Baca E, Villagomez-Castro JC, Mora-Montes HM, Flores-Carreon A, Toriello C.2011 г.
Sporothrix schenckii комплекс и споротрихоз, возникающая проблема со здоровьем. Будущий микробиол
6: 85–102. [PubMed] [Google Scholar] - Мареска Б., Кобаяши Г.С. 1989 г.
Диморфизм в Histoplasma capsulatum : модель для изучения дифференцировки клеток патогенных грибов. Microbiol Rev
53: 186–209. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Мареска Б., Кобаяши Г.С. 2000 г.
Диморфизм Histoplasma capsulatum и Blastomyces dermatitidis .Contrib Microbiol
5: 201–216. [PubMed] [Google Scholar] - Мареска Б., Каррату Л., Кобаяши Г.С. 1994 г.
Морфологический переход грибкового патогена человека Histoplasma capsulatum . Тенденции Microbiol
2: 110–114. [PubMed] [Google Scholar] - Мэрион К.Л., Рэппли, Калифорния, Энгл Дж. Т., Голдман МЫ. 2006 г.
Α- (1,4) -амилаза необходима для продукции α- (1,3) -глюканов и вирулентности в Histoplasma capsulatum . Мол Микробиол
62: 970–983. [PubMed] [Google Scholar] - Marsden-Haug N, Goldoft M, Ralston C, Limaye AP, Chua J, Hill H, Jecha L, Thompson GR III, Chiller T.2013.
Кокцидиоидомикоз приобретен в штате Вашингтон. Clin Infect Dis
56: 847–850. [PubMed] [Google Scholar] - Matute DR, McEwen JG, Puccia R, Montes BA, San-Blas G, Bagagli E, Rauscher JT, Restrepo A, Morais F, Nino-Vega G, et al.
2006a.
Тайное видообразование и рекомбинация у гриба Paracoccidioides brasiliensis , выявленные генеалогией. Мол Биол Эвол
23: 65–73. [PubMed] [Google Scholar] - Матуте Д. Р., Сепульведа В. Э., Кесада Л. М., Голдман Г. Х., Тейлор Дж. В., Рестрепо А., МакИвен Дж. Дж.2006b.
Микросателлитный анализ трех филогенетических видов Paracoccidioides brasiliensis . J Clin Microbiol
44: 2153–2157. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Мендес-Джаннини MJ, Монтейру да Силва JL, де Фатима да Силва J, Donofrio FC, Miranda ET, Андреотти PF, Soares CP. 2008 г.
Взаимодействие Paracoccidioides brasiliensis с клетками-хозяевами: последние достижения. Микопатология
165: 237–248. [PubMed] [Google Scholar] - Монтейро Дж. П., Клемонс К. В., Мирельс Л. Ф., Коллер Дж. А. младший, Ву Т. Д., Шанкар Дж., Лопес К. Р., Стивенс Д. А..2009 г.
Геномная микроматрица ДНК, сравнение паттернов экспрессии генов в мицелии Paracoccidioides brasiliensis и дрожжах in vitro. Микробиология
155: 2795–2808. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Neafsey DE, Barker BM, Sharpton TJ, Stajich JE, Park DJ, Whiston E, Hung CY, McMahan C, White J, Sykes S и др.
2010 г.
Популяционное геномное секвенирование грибов Coccidioides выявило недавнюю гибридизацию и контроль транспозонов. Genome Res
20: 938–946. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Nemecek JC, Wuthrich M, Klein BS.2006 г.
Глобальный контроль диморфизма и вирулентности грибов. Наука
312: 583–588. [PubMed] [Google Scholar] - Newman SL. 1999 г.
Макрофаги в защите хозяина от Histoplasma capsulatum . Тенденции Microbiol
7: 67–71. [PubMed] [Google Scholar] - Nguyen VQ, Sil A. 2008.
Индуцированное температурой переключение на патогенную дрожжевую форму Histoplasma capsulatum требует Ryp1, консервативного регулятора транскрипции. Proc Natl Acad Sci
105: 4880–4885. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Nguyen C, Barker BM, Hoover S, Nix DE, Ampel NM, Frelinger JA, Orbach MJ, Galgiani JN.2013.
Последние достижения в понимании экологических, эпидемиологических, иммунологических и клинических аспектов кокцидиоидомикоза. Clin Microbiol Rev
26: 505–525. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Nunes LR, Costa de Oliveira R, Leite DB, da Silva VS, dos Reis Marques E, da Silva Ferreira ME, Ribeiro DC, de Souza Bernardes LA, Goldman MH, Пуччиа Р. и др.
2005 г.
Транскриптомный анализ клеток Paracoccidioides brasiliensis , претерпевающих переход мицелия в дрожжи.Эукариотическая клетка
4: 2115–2128. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Ophuls W, Moffitt HC. 1900 г.
Новая патогенная плесень (ранее описываемая как простейшее: Coccidioides immitis pyogenes ): предварительный отчет. Филадельфия Мед Дж.
5: 1471–1472. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Паппагианис Д. 1988.
Эпидемиология кокцидиоидомикоза. Curr Top Med Mycol
2: 199–238. [PubMed] [Google Scholar] - Паппагианис Д., Линдси С., Билл С., Уильямс П.1979 г.
Этническое происхождение и клиническое течение кокцидиоидомикоза. Am Rev Respir Dis
120: 959–961. [PubMed] [Google Scholar] - Pappas PG, Tellez I, Deep AE, Nolasco D, Holgado W., Bustamante B. 2000.
Споротрихоз в Перу: Описание области гиперэндемичности. Clin Infect Dis
30: 65–70. [PubMed] [Google Scholar] - Пасрича С., Пейн М., Кановас Д., Пейс Л., Нгаосуванкул Н., Борода С., Ошлак А., Смит Г. К., Чайяродж С. К., Бойс К. Дж. И др.
2013.
Специфические для клеточного типа профили транскрипции диморфного патогена Penicillium marneffei отражают различные репродуктивные, морфологические и экологические требования.G3 (Bethesda)
3: 1997–2014. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Patel JB, Batanghari JW, Goldman WE. 1998 г.
Исследование дрожжевой фазовой экспрессии гена CBP1 в Histoplasma capsulatum . J Бактериол
180: 1786–1792. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Pfaller MA, Diekema DJ. 2010 г.
Эпидемиология инвазивных микозов в Северной Америке. Crit Rev Microbiol
36: 1–53. [PubMed] [Google Scholar] - Pine L. 1960.
Морфологические и физиологические характеристики Histoplasma capsulatum.Чарльз Томас, Спрингфилд, штат Иллинойс. [Google Scholar] - Посадас А. 1892.
Un nuevo caso de micosis Phydiodea con psorospermias [Новый случай грибкового микоза с псороспермией]. Энн Сирк Мед Арджент
15: 585–597. [Google Scholar] - Rappleye CA, Engle JT, Goldman WE. 2004 г.
РНК-интерференция в Histoplasma capsulatum демонстрирует роль α- (1,3) -глюкана в вирулентности. Мол Микробиол
53: 153–165. [PubMed] [Google Scholar] - Ричини-Перейра В.Б., Боско Сде М., Гриезе Дж., Теодоро Р.С., Макорис С.А., да Силва Р.Дж., Баррозо Л., Таварес П.М., Занкопе-Оливейра Р.М., Багагли Э.2008 г.
Молекулярное обнаружение Paracoccidioides brasiliensis в убитых на дорогах диких животных. Мед Микол
46: 35–40. [PubMed] [Google Scholar] - Rimek D, Zimmermann T, Hartmann M, Prariyachatigul C, Kappe R. 1999.
Распространена инфекция Penicillium marneffei у ВИЧ-инфицированной женщины из Таиланда в Германии. Микозы
42: 25–28. [PubMed] [Google Scholar] - Риксфорд Э. 1894.
Случай протазоанового дерматита. В западные медицинские времена, Vol. 8
Д. Джонстон, Сакраменто, Калифорния.[Google Scholar] - Робледо М.А., Грейбилл Дж. Р., Аренс Дж., Рестрепо А., Друц Д. Д., Робледо М. 1982.
Защита хозяина от экспериментального паракокцидиоидомикоза. Am Rev Respir Dis
125: 563–567. [PubMed] [Google Scholar] - Самсон Р.А., Йилмаз Н., Хубракен Дж., Шперенбург Х., Зайферт К.А., Петерсон С.В., Варга Дж., Фрисвад Дж. 2011 г.
Филогения и номенклатура рода Talaromyces и таксонов, включенных в подрод Penicillium Biverticillium . Стад Микол
70: 159–183.[Бесплатная статья PMC] [PubMed] [Google Scholar] - Sasaki AA, Fernandes GF, Rodrigues AM, Lima FM, Marini MM, Dos SFL, de Melo Teixeira M, Felipe MS, da Silveira JF, de Camargo ZP. 2014 г.
Хромосомный полиморфизм в комплексе Sporothrix schenckii . PLoS ONE
9: e86819. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Schenck B. 1898.
На невосприимчивых подкожных абсцессах, вызванных грибком, возможно, связанным со споротрихиями. Булл Джона Хопкинса
9: 286–290. [Google Scholar] - Sebghati TS, Engle JT, Goldman WE.2000 г.
Внутриклеточный паразитизм Histoplasma capsulatum : вирулентность грибов и зависимость от кальция. Наука
290: 1368–1372. [PubMed] [Google Scholar] - Сегретен Г. 1959 г.
Penicillium marneffei n.sp., возбудитель микоза ретикулоэндотелиальной системы. Микопатология
11: 327–353. [PubMed] [Google Scholar] - Шекспир Н., Нили М. 2011.
Каталог статей (Чарльза) Брюса Чатвина, 1963–89. Бодлианская библиотека, Оксфордский университет, Оксфорд, Великобритания.[Google Scholar] - Sharpton TJ, Stajich JE, Rounsley SD, Gardner MJ, Wortman JR, Jordar VS, Maiti R, Kodira CD, Neafsey DE, Zeng Q, et al.
2009 г.
Сравнительный геномный анализ возбудителей грибковых заболеваний человека Coccidioides и их родственников. Genome Res
19: 1722–1731. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Сильва С.С., Паес Х.С., Соарес С.М., Фернандес Л., Фелипе М.С. 2008 г.
Понимание патобиологии Paracoccidioides brasiliensis на основе анализа транскриптома — достижения и перспективы.Микопатология
165: 249–258. [PubMed] [Google Scholar] - Smith JA, Kauffman CA. 2009 г.
Эндемические грибковые инфекции у пациентов, получающих терапию ингибитором фактора некроза опухоли-α. Наркотики
69: 1403–1415. [PubMed] [Google Scholar] - Smith JA, Kauffman CA. 2010 г.
Бластомикоз. Proc Am Thorac Soc
7: 173–180. [PubMed] [Google Scholar] - Стинберген Дж. Н., Носанчук Дж. Д., Маллиарис С. Д., Касадевалл А. 2004.
Взаимодействие Blastomyces dermatitidis , Sporothrix schenckii и Histoplasma capsulatum с Acanthamoeba castellanii .Заразить иммунную
72: 3478–3488. [Google Scholar] - Салливан Т.Д., Руни П.Дж., Кляйн Б.С. 2002 г.
Agrobacterium tumefaciens интегрирует ДНК-переносчик в отдельные хромосомные участки диморфных грибов и дает гомокариотическое потомство из многоядерных дрожжей. Эукариотическая клетка
1: 895–905. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Sun SH, Huppert M, Vukovich KR. 1976 г.
Быстрая конверсия in vitro и идентификация Coccidioides immitis . J Clin Microbiol
3: 186–190.[Бесплатная статья PMC] [PubMed] [Google Scholar] - Supparatpinyo K, Khamwan C, Baosoung V, Nelson KE, Sirisanthana T. 1994.
Распространена инфекция Penicillium marneffei в Юго-Восточной Азии. Ланцет
344: 110–113. [PubMed] [Google Scholar] - Таварес А.Х., Сильва С.С., Дантас А., Кампос Е.Г., Андраде Р.В., Мараньяо А.К., Бриджидо М.М., Пассос-Сильва Д.Г., Фачин А.Л., Тейшейра С.М. и др.
2007 г.
Ранний транскрипционный ответ Paracoccidioides brasiliensis при интернализации мышиными макрофагами.Микробы заражают
9: 583–590. [PubMed] [Google Scholar] - Ustianowski AP, Sieu TPM, Day JN. 2008 г.
Penicillium marneffei Инфекция ВИЧ. Curr Opin Infect Dis
21: 31–36. [PubMed] [Google Scholar] - Vanittanakom N, Mekaprateep M, Sriburee P, Vanittanakom P, Khanjanasthiti P. 1995.
Эффективность флотационного метода выделения Penicillium marneffei из засеянной почвы. J Med Vet Mycol
33: 271–273. [PubMed] [Google Scholar] - Vanittanakom N, Cooper CRJ, Fisher MC, Sirisanthana T.2006 г.
Penicillium marneffei инфекция и последние достижения в области эпидемиологии и молекулярной биологии. Clin Microbiol Rev
19: 95–110. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Viriyakosol S, Singhania A, Fierer J, Goldberg J, Kirkland TN, Woelk CH. 2013.
Экспрессия генов грибкового патогена человека Coccidioides immitis изменяется по мере того, как артроконидии дифференцируются в шарики и созревают. BMC Microbiol
13: 121. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Wang H, LeBert V, Hung CY, Galles K, Saijo S, Lin X, Cole GT, Klein BS, Wuthrich M.2014 г.
Рецепторы лектина С-типа по-разному индуцируют клетки Th27 и иммунитет вакцины к эндемическому микозу Северной Америки. J Immunol
192: 1107–1119. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Webster RH, Sil A. 2008.
Консервативные факторы Ryp2 и Ryp3 контролируют морфологию клеток и образование инфекционных спор грибкового патогена Histoplasma capsulatum . Proc Natl Acad Sci
105: 14573–14578. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Wernicke R. 1892.
Ueber einen Protozoenbefund bei Mycosis фунгоидный.Центрбл Бактериол
12: 859–861. [Google Scholar] - Wheat J. 1997.
Гистоплазмоз. Опыт во время вспышек в Индианаполисе и обзор литературы. Медицина (Балтимор)
76: 339–354. [PubMed] [Google Scholar] - Пшеница LJ, Slama TG, Eitzen HE, Kohler RB, French ML, Biesecker JL. 1981 г.
Крупная городская вспышка гистоплазмоза: клинические особенности. Энн Интерн Мед
94: 331–337. [PubMed] [Google Scholar] - Whiston E, Zhang Wise H, Sharpton TJ, Jui G, Cole GT, Taylor JW.2012 г.
Сравнительная транскриптомика сапробной и паразитической фаз роста у Coccidioides spp. PLoS ONE
7: e41034. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Woods JP. 2003 г.
Стук в правую дверь и создание уютного дома: Histoplasma capsulatum Внутриклеточный патогенез. Curr Opin Microbiol
6: 327–331. [PubMed] [Google Scholar] - Woods JP, Goldman WE. 1992 г.
Получение in vivo линейных плазмид с добавлением теломерных последовательностей с помощью Histoplasma capsulatum .Мол Микробиол
6: 3603–3610. [PubMed] [Google Scholar] - Woods JP, Goldman WE. 1993 г.
Автономная репликация чужеродной ДНК в Histoplasma capsulatum : роль нативных теломерных последовательностей. J Бактериол
175: 636–641. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Woods JP, Heinecke EL, Goldman WE. 1998a.
Электротрансформация и экспрессия бактериальных генов, кодирующих гигромицинфосфотрансферазу и β-галактозидазу, в патогенном грибке Histoplasma capsulatum .Заразить иммунную
66: 1697–1707. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Woods JP, Retallack DM, Heinecke EL, Goldman WE. 1998b.
Редкий гомологичный ген, нацеленный на Histoplasma capsulatum : Нарушение гена URA5 Hc путем аллельной замены. J Бактериол
180: 5135–5143. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Вутрих М., Герн Б., Хунг С.Й., Эрсланд К., Рокко Н., Пик-Джейкобс Дж., Галлес К., Филутович Х., Уорнер Т., Эванс М. и др.2011 г.
Для защиты от 3 системных микозов, эндемичных для Северной Америки, вакцинация требует наличия у мышей клеток Th27. J Clin Invest
121: 554–568. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Wuthrich M, Ersland K, Pick-Jacobs JC, Gern BH, Frye CA, Sullivan TD, Brennan MB, Filutowicz HI, O’Brien K, Korthauer KD, et al. .
2012 г.
Ограниченная модельная экспрессия антигена трансгенными грибами вызывает различные судьбы во время дифференцировки адоптивно перенесенных Т-клеточных рецепторов трансгенных CD4 + Т-клеток: сильная активация и пролиферация со слабой эффекторной функцией во время отзыва.Заразить иммунную
80: 787–797. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Youseff BH, Dougherty JA, Rappleye CA. 2009 г.
Обратная генетика посредством случайного мутагенеза в Histoplasma capsulatum . BMC Microbiol
9: 236. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Youseff BH, Holbrook ED, Smolnycki KA, Rappleye CA. 2012 г.
Внеклеточная супероксиддисмутаза защищает дрожжевые клетки Histoplasma от окислительного стресса, вызванного хозяином. PLoS Pathog
8: e1002713. [Бесплатная статья PMC] [PubMed] [Google Scholar] - Zhang Y, Li G, He D, Yu B, Yokoyama K, Wang L.2011 г.
Эффективная система инсерционного мутагенеза для диморфного патогенного гриба Sporothrix schenckii с использованием Agrobacterium tumefaciens . J Microbiol методы
84: 418–422. [PubMed] [Google Scholar]
Молекулярная биология и патогенные механизмы
Патогенные грибы человека: молекулярная биология и патогенные механизмы
«тщательно отредактированный и хорошо подготовленный справочник» (IMA Fungus)
«настоятельно рекомендуется … полезный источник информации» (грибковое разнообразие)
«ценный справочник для специалистов в этой области, а также тех, кто плохо знаком с грибковыми заболеваниями. патобиология »(Frontiers in Microbiology)
Издатель: Caister Academic Press
Отредактировал: Дерек Дж.Салливан и Гэри П. Моран
Отделение биологических наук о полости рта, Школа стоматологических наук и Больница стоматологического университета Дублина, Тринити-колледж Дублин, Дублин 2, Ирландия
Страницы: x + 342
Переплет:
Дата публикации: июнь 2014 г.
ISBN: 978- 1-0-44-7
Цена: 219 фунтов стерлингов или 360 долларов США Купить книгу или купить онлайн
Электронная книга:
Дата публикации: май 2014 г.
ISBN: 978-1-0-66-9
Цена: 360 долларов США Купить электронную книгу
DOI: https://doi.org/10.21775/97819101
Те, кто смотрели эту книгу, также смотрели:
Грибки вызывают у людей широкий спектр заболеваний, от сравнительно безобидных поверхностных кожных заболеваний, вызываемых дерматофитами, до инвазивных опасных для жизни инфекций, вызываемых такими видами, как Candida albicans, или Cryptococcus neoformans. Из-за условно-патогенной природы большинства инвазивных микозов грибковую патогенность определить трудно. Однако применение новых геномных и других молекулярных методов в последние годы произвело революцию в этой области, открыв захватывающие новые представления о механизмах грибкового патогенеза.
В этой книге группа известных авторов критически рассматривает наиболее важные исследования, чтобы обеспечить своевременный обзор. Обширные справочные разделы в каждой главе положительно побуждают читателей изучить предмет более подробно.Книга разделена на два раздела: в первых шести главах рассматривается преобразующий эффект применения современных инструментов и новаторских подходов к исследованиям, особенно в области сравнительной биологии. Второй раздел состоит из восьми глав, каждая из которых посвящена молекулярной и клеточной биологии основного грибкового патогена человека: Candida, Aspergillus, Cryptococcus, дерматофитов, Histoplasma, Blastomyces, Pneumocystis и Paracoccidoides .Эти главы представляют собой своевременную картину текущего состояния исследований.
Этот том является важным справочным материалом для студентов, исследователей и клиницистов, интересующихся грибковым патогенезом.
Отзывы
«Редакторам … удалось собрать вместе 34 самых активных и ведущих исследователя в этой области для создания такого обзора … цветные диаграммы и фотографии настолько информативны и (или) потрясающи, что я могу себе представить, что некоторые из них будут широко использоваться при преподавании курсов медицинской микологии… Это тщательно отредактированный и хорошо подготовленный справочник, который заслуживает широкого доступа в лабораториях, изучающих молекулярную биологию и механизмы патогенности патогенных грибов человека «из IMA Fungus (2014) 5: 56-57.
» эта книга настоятельно рекомендуется … должен быть доступен во всех библиотеках колледжей и университетов, где предлагаются курсы по патологии человека и биотехнологии. Это также полезный источник информации как для студентов, так и для исследователей. «Из разнообразия грибов
» Современные, хорошо представленные главы служат для того, чтобы сделать этот текст ценным справочным материалом для специалистов в данной области, а также для тех, кто новое в грибковой патобиологии…. эта хорошо представленная работа является очень полезным дополнением к литературе … Чрезвычайно удобочитаемый характер хорошо отредактированного текста, безусловно, облегчит его использование аспирантами, занимающимися фундаментальной или медицинской микологией, и признанные исследователи оценят четкие обзоры и подробные ссылки на них. «от Frontiers in Microbiology
Содержание
1. Понимание патогенеза грибков с помощью высокопроизводительного секвенирования
Винсент Бруно
Изобретение массового параллельного секвенирования ДНК и последующее развитие секвенирования RNA-seq, ChIP-seq и микробиома радикально изменили ландшафт молекулярной и системной биологии. позволяя исследователям решать биологические проблемы с такой чувствительностью и масштабом, которые ранее были невозможны.В этой главе обсуждается, как каждый из этих подходов, основанных на высокопроизводительном секвенировании (HTS), используется для решения биологических вопросов в области патогенеза грибков у человека. По мере того, как стоимость секвенирования становится более доступной, достижимая степень охвата глубины последовательности значительно возрастет, что позволит исследователям применить эти геномные методы к большему количеству моделей in vivo и клинических образцов, для которых требуется большой охват. Интегрированные подходы, сочетающие несколько экспериментов на основе HTS, обеспечат необходимое понимание сложных взаимодействий между хозяином, микробиотой и грибковыми патогенами.
2. Сравнительная геномика и эволюционный анализ грибковых патогенов человека
Дэвид А. Фитцпатрик
Недавние достижения в методах секвенирования следующего поколения резко увеличили доступность геномных данных. Из-за их относительно небольшого размера генома и важности их в качестве патогенов человека и сельскохозяйственных культур на сегодняшний день было полностью секвенировано и опубликовано более сотни геномов грибов. Ожидается, что это число резко возрастет в ближайшие годы, когда отдельные учреждения будут секвенировать свои собственные представляющие интерес грибы и инициировать амбициозный проект по секвенированию 1000 забытых геномов грибов (https: // 1000.фунгалгеномес. орг /). В настоящее время значительную часть имеющихся геномов составляют патогены человека, а также близкородственные непатогенные виды. Этот тип данных позволяет нам проводить сравнительный геномный анализ патогенных и непатогенных видов в попытке раскрыть молекулярные механизмы, связанные с заболеванием. Наличие полных последовательностей генома также позволяет нам исследовать многие эволюционные процессы в грибах, такие как горизонтальный перенос генов, слияние / деление генов, а также прирост и потеря интронов.В следующей главе будет обсужден выбор ключевых сравнительных геномных анализов, которые были выполнены на ряде наиболее распространенных патогенов человека, а также показано, как полные геномы грибов помогли нам понять некоторые механизмы, которые сформировали эволюцию грибов.
3. Системная биология грибковых инфекций, управляемая данными
Фабиан Хорн, Вито Валианте, Рейнхард Гутке и Аксель А. Брахаге
Последние технические достижения в биотехнологии открыли новый захватывающий период исследований.Высокопроизводительные данные могут быть получены в рамках биомолекулярных экспериментов с разумными усилиями и затратами. Это позволяет исследовать взаимодействия хозяин-грибок на общесистемном уровне. Узкое место в рабочем процессе эксперимента сместилось от получения данных к их надлежащему анализу. Анализ данных с высокой пропускной способностью можно описать операционным протоколом системной биологии, управляемой данными. Протокол включает экспериментальный план, предварительную обработку данных, выбор функций, компьютерное моделирование и биологическую оценку.В ходе этой процедуры данные необходимо анализировать в соответствующем биологическом контексте. Цель конвейера анализа — получить переменные и параметры модели, чтобы сделать новые гипотезы для дальнейших экспериментов. Переплетенный характер взаимодействия хозяин-патоген также отражается в комплексном анализе данных, в ходе которого в настоящее время собирается первый опыт и разрабатываются новые методы. Актуальными проблемами являются интеграция различных уровней омикс-данных, т.е. моделирование должно учитывать несколько молекулярных уровней и интеграцию различных технологий на каждом уровне.Ожидается, что достижения в системной биологии помогут перевести знаний in vivo о грибковых инфекциях в клинические приложения.
4. Сравнительный патогенез: транскриптомный анализ взаимодействий клетки-хозяина и грибка
Элейн Бигнелл
Вклад активности хозяина в положительные и отрицательные исходы грибковых заболеваний становится все более очевидным. Технологические достижения позволяют ученым по отдельности разгадывать сложности взаимодействия хозяина и патогена — цель, которая останется важнейшим компонентом исследований инфекционных заболеваний в обозримом будущем и, вероятно, приведет к появлению нового поколения иммунотерапевтических противогрибковых организмов.Взаимодействие грибка-хозяина-патогена включает в себя замечательный набор поведений, от опосредованного патогеном уничтожения клеток-хозяев до внутриклеточного заселения и литического изгнания, поведения, которое определяет исход болезни, управляя репликацией патогена, уклонением от иммунитета, выживанием и распространением. В этой главе будут изучены результаты транскриптомного анализа взаимодействий грибов-хозяин-патоген.
5. Животные модели грибковой инфекции человека
Donna M. MacCallum
Грибковая инфекция является основной причиной заболеваемости и смертности, причем большинство инфекций вызываются дерматофитами, Candida видов , Aspergillus видов, Cryptococcus neoformans и Pneumocystis jirovecii .Хотя дерматофиты могут считаться первичными патогенами, большинство грибковых инфекций вызывается условно-патогенными видами грибов. Инфекции Candida , как правило, происходят из комменсальных источников, но инфекции Aspergillus , Cryptococcus и Pneumocystis передаются из окружающей среды. Эти инфекции обнаруживаются у тяжелобольных пациентов или у лиц с ослабленным иммунитетом, особенно у людей с ВИЧ / СПИДом. Смертность, связанная с этими инфекциями, остается высокой, частично из-за трудностей в диагностике, но также из-за ограниченного выбора терапевтических средств.Чтобы облегчить разработку этих крайне необходимых диагностических и противогрибковых средств, требуется более глубокое понимание грибкового патогенеза. Модели на животных дают возможность исследовать начало и прогрессирование грибковых заболеваний, а также оценить новые противогрибковые препараты. Описаны животные модели инфекций дерматофитов, Candida , Aspergillus , Cryptococcus и Pneumocystis , и обсужден их вклад в наше текущее понимание этих важных грибковых инфекций.
6. Ответы хозяина на грибковую инфекцию
Дэвид Л. Мойес, Джонатан П. Ричардсон и Джулиан Р. Наглик
Три основных условно-патогенных грибковых патогена человека — это Candida albicans , Aspergillus fumigatus и Cryptococcus neoformans. Эти патогены в первую очередь инфицируют людей с ослабленным иммунитетом либо в результате иммунодефицита (например, нейтропения, ВИЧ-инфекция), либо в результате интервенционной терапии (например, лейкемии, трансплантации органов, рака).Это подчеркивает важность иммунной защиты хозяина в контроле или предотвращении грибковых инфекций. В здоровом организме эти грибковые патогены первоначально взаимодействуют с поверхностями слизистой оболочки, что может привести к колонизации и установлению комменсализма, но у людей с ослабленным иммунитетом эти грибки могут вызывать серьезные поверхностные инфекции и могут распространяться, вызывая опасные для жизни системные инфекции. Для борьбы с этими грибковыми инфекциями хозяин использует обширную коммуникационную сеть клеток, белков и химических сигналов, распространяемых в крови и тканях, которые составляют врожденный и адаптивный иммунитет.В этой главе мы рассмотрим, как хозяин распознает эти грибы, события, вызванные грибковыми клетками, и иммунную защиту хозяина, которая в конечном итоге устраняет инфекции во время выздоровления. Обзор будет в первую очередь нацелен на C. albicans , наиболее распространенный грибковый патоген человека.
7. Молекулярное обновление патогенеза кандидоза
Дункан Уилсон и Бернхард Хьюбе
В отличие от большинства патогенных грибов человека, описанных в этой книге, патогенные виды рода Candida обычно не существуют в экологических нишах, таких как в виде почвы или компоста.Вместо этого эти грибы развивались в тесной связи с теплокровными животными, такими как люди. Для большинства людей эти дрожжи в большинстве случаев существуют как безвредные члены микробиома. Однако некоторые виды способны вызывать разрушительные инфекции у своих хозяев-людей. В этой главе мы обобщаем и обсуждаем важные недавние изменения, которые были сделаны в нашем понимании молекулярной основы патогенности Candida , с акцентом на доминирующий патогенный вид, Candida albicans .
8. Характеристики вирулентности Aspergillus fumigatus
Ребекка А. Оуэнс, Грейн О’Киф, Карен А. О’Ханлон, Лорна Галлахер и Шон Дойл
Aspergillus fumigatus является условно-патогенным микроорганизмом, вызывающим серьезное грибковое заболевание. широко называемый аспергиллез у иммунокомпетентных и иммунодефицитных лиц. Достижения в диагностике аспергиллеза основаны на обнаружении нуклеиновой кислоты грибов с помощью ПЦР и секретируемого галактоманнана с помощью ELISA.Появляются новые диагностические стратегии, основанные на обнаружении секретируемых низкомолекулярных метаболитов. Для лечения инфекции, вызванной A. fumigatus , доступны лекарственные препараты, которые нарушают биосинтез и целостность мембран, а также функцию клеточных стенок. По-видимому, нет единственной характеристики, обеспечивающей полную вирулентность A. fumigatus , скорее, она, по-видимому, зависит от комбинации факторов выживания, частично обусловленных его функцией сапрофита, для облегчения колонизации хозяином и роста грибов.Эти атрибуты включают прочную клеточную стенку, небольшие гидрофобные конидии и архитектуру биопленок; устойчивость к окислительному стрессу, гипоксии и железному голоданию; биосинтез и секреция вторичных метаболитов, а также термотолерантность и способность секретировать деструктивные ферменты для поглощения питательных веществ. Снижение эффективности этих стратегий выживания открывает возможности для разработки новых классов противогрибковых препаратов.
9. Cryptococcus
Росио Гарсия-Родас, Радамес Дж.Б. Кордеро и Оскар Сарагоса
Cryptococcus neoformans — космополитический патоген, вызывающий серьезную озабоченность среди ВИЧ-инфицированных пациентов и широко распространенный в развивающихся странах. Cryptococcus neoformans При вдыхании бластококков или высохших спор они могут вызвать пневмонию и менингит у лиц с иммунодефицитом, что приводит к летальному исходу, если не лечить. Его родственный вид Cryptococcus gattii более ограничен тропическими зонами и поражает в основном иммунокомпетентных людей. Взаимодействия Cryptococcus с человеческим хозяином были широко изучены, в частности его способность модулировать иммунный ответ, реплицироваться в макрофагах, распространяться из легких в центральную нервную систему и / или оставаться в латентном состоянии в течение многих лет. В этой главе будет рассмотрено наше текущее понимание детерминант вирулентности, которые способствуют возникновению инфекции Cryptococcus , в частности его полисахаридная капсула и накопление меланина, а также другие черты, которые придают адаптации хозяину.
10. Дерматофиты как сапрофиты и патогены
Мишель Моно, Бернар Миньон и Питер Стейб
Дерматофиты поражают роговой слой , ногти и волосы и являются наиболее распространенными возбудителями поверхностных микозов у людей и животных. В настоящее время секвенирован геном семи видов. Размер геномов дерматофитов от 22,5 до 24 МБ меньше, чем у Coccidioides spp., Histoplasma spp. и Aspergillus spp.Они обогащены определенными семействами генов, кодирующих секретируемые протеазы, специфические киназы грибов и белки, содержащие домен LysM, который, как известно, связывает хитин. Недавно были разработаны различные инструменты для улучшения генетического анализа дерматофитов, включая эффективные системы для целевой инактивации генов, подавления генов и широкие методы транскрипционного профилирования. Неожиданно было обнаружено, что профили экспрессии генов в коже и волосах инфицированных морских свинок сильно отличаются от профилей во время роста in vitro с использованием твердого кератина в качестве субстрата.Было обнаружено, что вместо основных генов протеазы, экспрессируемых in vitro , другие активируются только в коже инфицированных животных. Другими словами, экспрессия предполагаемых генов вирулентности при дерматофитозах более сложна, чем предполагалось ранее, и, вероятно, зависит от места и типа инфекции. Дальнейшие подходы к широкому транскрипционному профилированию во время инфекций дадут новое понимание патогенных механизмов дерматофитов.
11. Механизмы патогенеза Histoplasma capsulatum
Chad A.Rappleye
Диморфный грибковый патоген Histoplasma capsulatum вызывает респираторные и системные заболевания как у лиц с ослабленным иммунитетом, так и у иммунокомпетентных людей. У млекопитающих-хозяев Histoplasma растет как патогенные дрожжи, которые выживают и размножаются внутри фагоцитов. Генетические исследования идентифицировали сигнальную киназу и 3 фактора транскрипционных факторов, которые регулируют термически индуцированный процесс дифференцировки дрожжевой фазы. Механизмы, которые способствуют патогенезу Histoplasma (суммировано на рисунке 11.1) сосредоточены на трех основных задачах, стоящих перед внутриклеточными патогенами: (1) сокрытие молекулярных сигнатур, чтобы избежать обнаружения фагоцитами и активации иммунной защиты, (2) детоксикация молекул антимикробной защиты, продуцируемых фагоцитами, и (3) получение необходимых питательных веществ для рост и репликация внутри фагосомы, ограниченной питательными веществами. Доказанные факторы вирулентности, обеспечивающие эти необходимые аспекты патогенеза, включают синтез α-глюкана, который маскирует иммуностимулирующие β-глюканы клеточной стенки дрожжей, выработку и секрецию молекул антиоксидантов (внеклеточной супероксиддисмутазы и каталазы) для специфической защиты от реактивного кислорода, происходящего из фагоцитов, и секреции множества факторов приобретения железа (сидерофоры и железоредуктазы).Кроме того, дрожжи Histoplasma секретируют несколько новых факторов с неопределенными функциями, но которые, вероятно, связаны с патогенезом в силу их специфической экспрессии только клетками патогенной фазы и их внеклеточной локализации. Вместе эти механизмы позволяют дрожжам Histoplasma успешно создавать инфекции, несмотря на полностью функциональную иммунную защиту хозяина.
12. Blastomyces dermatitidis и Blastomycosis
Грегори М.Gauthier
Этиологический агент бластомикоза, Blastomyces dermatitidis , принадлежит к группе грибов аскомицетов, проявляющих термический диморфизм. В почве (22-25 ° C) эти грибы растут в виде нитчатой плесени, вызывающей инфекционные конидии. После разрушения почвы такими видами деятельности, как строительство, аэрозольные конидии и фрагменты плесени попадают в легкие хозяина (37 ° C) и превращаются в патогенные дрожжи. В дрожжевой форме B. dermatitidis способен уклоняться от иммунной защиты, изменяя состав клеточной стенки грибов и нарушая цитокиновый ответ хозяина.Примерно у 25-40% пациентов, инфицированных B. dermatitidis , развивается диссеминированное заболевание, которое часто поражает кожу или кости. Диагностика бластомикоза требует высокой степени клинического подозрения в сочетании с использованием культуральных и некультуральных диагностических средств. Лечение требует использования полиеновых или азольных противогрибковых средств.
13. Новые сведения о патогенезе Pneumocystis Pneumonia
Jakrapun Pupaiboo и Andrew H. Limper
Pneumocystis пневмония (PCP) остается основной причиной заболеваемости и смертности среди ВИЧ-инфицированных пациентов.Кроме того, распространенность PCP увеличивается у пациентов, не инфицированных ВИЧ, с подавленным иммунным статусом после использования мощных иммуносупрессивных методов лечения. Наше понимание Pneumocystis основано на моделях животных с подавленным иммунитетом. Исследования жизненного цикла, патогенеза и иммунного ответа хозяина Pneumocystis представляют собой привлекательные мишени для разработки новых агентов против Pneumocystis . Сульфаметоксазол-триметоприм остается схемой первой линии для лечения и профилактики PCP.Многие аспекты биологии Pneumocystis остаются без ответа или спорными, такие как респираторная колонизация, способ передачи, клиническое значение мутаций в доступных в настоящее время агентах против Pneumocystis , а также профилактика и лечение пациентов с иммунодефицитом, не связанным с ВИЧ. Существует острая необходимость в продолжении исследований этого уникального патогенного грибка, важного с медицинской точки зрения, чтобы мы могли разработать более эффективные стратегии профилактики и лечения инфекции Pneumocystis .
14. Paracoccidioides Механизмы патогенеза и вирулентности
Juliana Alves Parente, Clayton Luiz Borges, Maristela Pereira, Alexandre Melo Bailão, Rosely Maria Zancopé Oliveira и Célia Maria de Almeida Soares
63 Возбудителя Paracusocus 916 являются паразитами 9. возбудители одного из наиболее распространенных системных микозов в Латинской Америке, а именно паракокцидиоидомикоза (ПКМ). Исследования сообщили о разной степени патогенности и вирулентности среди представителей этого рода и предоставили доказательства вариабельности восприимчивости хозяина к инфекции.Идентификации и валидации факторов вирулентности препятствовали трудности в генетической манипуляции с этим патогеном, хотя исследования показали, что патогенность Paracoccidioides обеспечивается рядом признаков вирулентности, включая диморфизм грибов, состав клеточной поверхности и адгезию. для тканей хозяина секреция молекул, составляющих арсенал вирулентности гриба, конкуренция с хозяином за основные питательные вещества, такие как железо, и последующая адаптация патогена к окружающей среде хозяина.В этой главе представлена информация о современных знаниях о вирулентности рода Paracoccidioides .
Как купить эту книгу
(EAN: 97810447 97810669 Субъекты: [микробиология] [геномика] [микология])
Обзор грибковых инфекций — инфекционные заболевания
Серологические тесты (в основном для Aspergillus , Blastomyces , Candida , Coccidioides , Cryptococcus и Histoplasma )
Если врачи подозревают острую или хроническую первичную грибковую инфекцию, они должны получить подробную историю поездок и проживания, чтобы определить, могли ли пациенты подвергаться воздействию определенных эндемических микозов, возможно, несколько лет назад.
Легочные грибковые инфекции следует отличать от опухолей и хронических пневмоний, вызываемых негрибковыми организмами, такими как микобактерии (включая туберкулез). Берут образцы для культивирования грибов и микобактерий и гистопатологии. Образцов мокроты может быть достаточно, но иногда может потребоваться бронхоальвеолярный лаваж, трансторакальная пункционная биопсия или даже хирургическое вмешательство для получения приемлемого образца.
Грибки, вызывающие первичные системные инфекции, легко распознать по их гистопатологическому внешнему виду.Однако идентификация конкретного гриба может быть трудной и обычно требует грибковой культуры. Клиническое значение положительных культур мокроты может быть неясным, если они показывают комменсальные организмы (например, Candida albicans ) или грибы, повсеместно распространенные в окружающей среде (например, Aspergillus видов). Следовательно, для постановки диагноза могут потребоваться другие доказательства (например, факторы хозяина, такие как иммуносупрессия, серологические данные, тканевая инвазия).
Серологические тесты можно использовать для проверки многих системных микозов, если культуральные и гистопатологические исследования недоступны или не выявлены, хотя немногие из них позволяют установить окончательный диагноз.К особенно полезным тестам относятся следующие:
Измерение специфических для организма антигенов, в первую очередь видов Cryptococcus neoformans , Histoplasma capsulatum и Aspergillus (в каждом из этих серологических тестов была отмечена случайная перекрестная реактивность с другими грибами)
Сывороточный бета-глюкан, который часто бывает положительным при инвазивном кандидозе, а также Pneumocystis jirovecii инфекциях
Анализы фиксации комплемента и новейшие иммуноферментные анализы на антикокцидиоидные антитела, которые удовлетворительно специфичны и не требуют подтверждения повышения уровня (высокие титры подтверждают диагноз и указывают на высокий риск внелегочного распространения)
Большинство других тестов на противогрибковые антитела имеют низкую чувствительность, специфичность или и то, и другое, и, поскольку требуется измерение титров в остром периоде и в период выздоровления, не могут использоваться для определения начальной терапии.
Молекулярная диагностика — новые полезные инструменты для идентификации молекулярных компонентов некоторых грибковых инфекций. Утвержденные FDA тесты включают ДНК-зонды, в которых используются образцы культур для идентификации Histoplasma , Blastomyces и Coccidioides , а также тесты полимеразной цепной реакции или ДНК-гибридизации, в которых используются образцы культур крови для идентификации Candida . Матричная лазерная десорбционная ионизация-времяпролетная масс-спектрометрия (MALDI-TOF), проведенная на образцах культур, также может быть использована для идентификации видов Candida .
14.6A: Грибы — Biology LibreTexts
Гриб является членом большой группы эукариотических организмов, в которую входят микроорганизмы, проявляющие патогенность.
Ключевые термины
- симбионт : организм, живущий в симбиотических отношениях; симбиот.
- микотоксин : любое вещество, вырабатываемое плесенью или грибком, которое является вредным для позвоночных при проглатывании, вдыхании или контакте с кожей.
- оппортунист : когда организм использует любую возможность для улучшения своей ситуации.
Грибок является членом большой группы эукариотических организмов, в которую входят такие микроорганизмы, как дрожжи и плесень. Эти организмы классифицируются как царство грибов, отдельно от растений, животных и бактерий. Грибы распространены по всему миру и могут расти в самых разных средах обитания, включая экстремальные условия, такие как пустыни или районы с высокой концентрацией соли или ионизирующей радиации, а также в глубоководных отложениях.Большинство грибов незаметны из-за небольшого размера их структур и загадочного образа жизни в почве, на мертвом веществе и в качестве симбионтов растений, животных или других грибов. Грибы играют важную роль в разложении органических веществ и играют фундаментальную роль в круговороте и обмене питательных веществ. Многие виды грибов производят биоактивные соединения, называемые микотоксинами, такие как алкалоиды и поликетиды, которые токсичны для животных, включая человека, способствуя патогенности и болезням.
Изучение патогенных грибов относится к медицинской микологии. Существуют различные примеры патогенных грибов, включая, помимо прочего, следующие виды: Candida, Aspergillosis, Cryptococcus, Histoplasma, Pneumocystis и Stachybotrys.
Обычно известно, что виды Candida вызывают оппортунистические инфекции у хозяев с ослабленным иммунитетом. Хозяева с ослабленным иммунитетом, которые обычно заражаются Candida, включают пациентов после трансплантации, больных раком и больных СПИДом. Инфекции Candida трудно поддаются лечению и могут привести к системным инфекциям, ведущим к смерти.
Рисунок: Candida : Candida-инфекция, обнаруженная на образце Папаниколау.
Один из наиболее распространенных грибковых патогенных видов включает штаммы Aspergillus, в частности Aspergillus fumigatus и Aspergillus flavus. Aspergillus может вызывать заболевание через выработку микотоксинов, индукцию аллергических реакций и через локальные или системные инфекции. Aspergillus flavus специфически продуцирует афлатоксин, который одновременно является токсином и канцерогеном, тогда как Aspergillus fumigatus вызывает аллергическое заболевание.Симптомы заболеваний, вызываемых Aspergillus, могут включать жар, кашель, боль в груди или одышку.
Cryptococcus neoformans вызывает тяжелые формы менингита и менингоэнцефалита у пациентов с ВИЧ-инфекцией и СПИДом. Виды Cryptococcus живут в почве и не вызывают заболеваний у людей, поэтому Cryptococcus neoformans является основным патогеном как для человека, так и для животных.
Histoplasma capsulatum вызывает образование гистоплазмоза у людей, собак и кошек. Этот специфический грибок является эндемическим в некоторых районах США, и заражение происходит при вдыхании загрязненного воздуха.
Pneumocystis jirovecii приводит к развитию пневмонии у людей с ослабленной иммунной системой, включая недоношенных детей, пожилых людей и больных СПИДом.
Stachybotrys chartarum, также называемый черной плесенью, вызывает поражение органов дыхания и сильные головные боли. Этот тип черной плесени часто встречается в семьях с хронической влажностью.
грибов между крайностью толерантности и условно-патогенной патогенностью для человека
Ахмед С.А., ван де Санде WWJ, Стивенс Д.А., Фахал А., ван Дипенинген А.Д., Менкен SBJ, де Хуг Г.С. (2014) Пересмотр возбудителей чернозерной эвмицетомы в порядке Pleosporales.Персона 33: 141–154. https://doi.org/10.3767/003158514×684744
CAS
Статья
PubMed
PubMed Central
Google Scholar
Al-Hatmi AMS, Meis JF, de Hoog GS (2016) Fusarium: молекулярное разнообразие и внутренняя лекарственная устойчивость. PLoS Pathog. https://doi.org/10.1371/journal.ppat.1005464
Артикул
PubMed
PubMed Central
Google Scholar
Amselem J, Cuomo CA, van Kan JAL, Viaud M, Benito EP, Couloux A, Coutinho PM, de Vries RP, Dyer PS, Fillinger S, Fournier E, Gout L, Hahn M, Kohn L, Lapalu N , Plummer KM, Pradier JM, Quevillon E, Sharon A, Simon A, ten Have A, Tudzynski B, Tudzynski P, Wincker P, Andrew M, Anthouard V, Beever RE, Beffa R, Benoit I, Bouzid O, Brault B, Chen ZH, Choquer M, Collemare J, Cotton P, Danchin EG, Da Silva C, Gautier A, Giraud C, Giraud T, Gonzalez C, Grossetete S, Guldener U, Henrissat B, Howlett BJ, Kodira C, Kretschmer M, Lappartient A, Leroch M, Levis C, Mauceli E, Neuveglise C, Oeser B, Pearson M, Poulain J, Poussereau N, Quesneville H, Rascle C, Schumacher J, Segurens B, Sexton A, Silva E, Sirven C, Soanes DM, Talbot NJ, Templeton M, Yandava C, Yarden O, Zeng QD, Rollins JA, Lebrun MH, Dickman M (2011) Геномный анализ патогенов экротрофных грибов Sclerotinia sclerotiorum и Botrytis cinerea .PLoS Genet. https://doi.org/10.1371/journal.pgen.1002230
Артикул
PubMed
PubMed Central
Google Scholar
Армийос Харамилло В.Д., Варгас В.А., Сукно С.А., Тон М.Р. (2013) Горизонтальный перенос гена субтилизина от растений к предку растительного патогенного грибка рода Colletotrichum. PLoS ONE 8 (3): e59078. https://doi.org/10.1371/journal.pone.0059078
CAS
Статья
PubMed
PubMed Central
Google Scholar
Arnaud MB, Chibucos MC, Costanzo MC, Crabtree J, Inglis DO, Lotia A, Orvis J, Shah P, Skrzypek MS, Binkley G, Miyasato SR, Wortman JR, Sherlock G (2010) База данных генома Aspergillus , кураторский ресурс по сравнительной геномике для информации о генах, белках и последовательностях для исследовательского сообщества Aspergillus .Nucleic Acids Res 38: 420–427. https://doi.org/10.1093/Nar/Gkp751
Артикул
Google Scholar
Baker RM, Stegink RJ, Manaloor JJ, Schmitt BH, Stevens JC, Christenson JC (2016) Malassezia pneumonia : редкое осложнение терапии парентеральным питанием. JPEN J Parenter Enteral Nutr 40 (8): 1194–1196. https://doi.org/10.1177/0148607115595224
Артикул
PubMed
Google Scholar
Blin K, Medema MH, Kazempour D, Fischbach MA, Breitling R, Takano E, Weber T (2013) antiSMASH 2.0 — универсальная платформа для анализа генома продуцентов вторичных метаболитов. Nucleic Acids Res 41 (1): 204–212. https://doi.org/10.1093/nar/gkt449
Артикул
Google Scholar
Brown SM, Campbell LT, Lodge JK (2007) Cryptococcus neoformans , грибок в состоянии стресса. Curr Opin Microbiol 10 (4): 320–325. https://doi.org/10.1016/j.mib.2007.05.014
CAS
Статья
PubMed
PubMed Central
Google Scholar
Bushley KE, Turgeon BG (2010) Филогеномика выявляет подсемейства грибковых нерибосомных пептидных синтетаз и их эволюционные отношения.BMC Evol Biol. https://doi.org/10.1186/1471-2148-10-26
Артикул
PubMed
PubMed Central
Google Scholar
Casadevall A (2007) Детерминанты вирулентности патогенных грибов. Fungal Biol Rev 21 (4): 130–132. https://doi.org/10.1016/j.fbr.2007.02.007
Артикул
PubMed
PubMed Central
Google Scholar
Casadevall A, Pirofski LA (2007) Случайная вирулентность, скрытый патогенез, марсиане, потерянные хозяева и патогенность микробов окружающей среды.Эукариотическая клетка 6 (12): 2169–2174. https://doi.org/10.1128/Ec.00308-07
CAS
Статья
PubMed
PubMed Central
Google Scholar
Casadevall A, Pirofski LA (2014) Микробиология: откажитесь от термина «патоген». Nature 516 (7530): 165–166. https://doi.org/10.1038/516165a
CAS
Статья
PubMed
Google Scholar
Casadevall A, Fang FC, Pirofski LA (2011) Вирулентность микробов как эмерджентное свойство: последствия и возможности.PLoS Pathog. https://doi.org/10.1371/journal.ppat.1002136
Артикул
PubMed
PubMed Central
Google Scholar
Cerqueira GC, Arnaud MB, Inglis DO, Skrzypek MS, Binkley G, Simison M, Miyasato SR, Binkley J, Orvis J, Shah P, Wymore F, Sherlock G, Wortman JR (2014) База данных генома Aspergillus: мультивидовое курирование и включение данных RNA-Seq для улучшения структурных аннотаций генов.Nucleic Acids Res 42 (D1): 705-710. https://doi.org/10.1093/nar/gkt1029
CAS
Статья
Google Scholar
Chen ZH, Martinez DA, Gujja S, Sykes SM, Zeng QD, Szaniszlo PJ, Wang Z, Cuomo CA (2014) Сравнительный геномный и транскриптомный анализ Wangiella dermatitidis , основной причины феогифомикоза и модельного черного дрожжевой возбудитель человека. G3 Genom Genet 4 (4): 561–578. https: // doi.org / 10.1534 / g3.113.009241
CAS
Статья
Google Scholar
Chowdhary A, Perfect J, de Hoog GS (2015) Черная плесень и меланизированные дрожжи, патогенные для человека. Cold Spring Harb Perspect Med. https://doi.org/10.1101/cshperspect.a019570
Артикул
PubMed Central
Google Scholar
Condon BJ, Ленг YQ, Wu DL, Bushley KE, Ohm RA, Otillar R, Martin J, Schackwitz W, Grimwood J, MohdZainudin N, Xue CS, Wang R, Manning VA, Dhillon B, Tu ZJ, Steffenson Б.Дж., Саламов А., Сан Х., Лоури С., ЛаБутти К., Хан Дж., Коупленд А., Линдквист Е., Барри К., Шмутц Дж., Бейкер С.Е., Чуффетти Л.М., Григорьев И.В., Чжун С., Тургеон Б.Г. (2013) Сравнительная структура генома, вторичный метаболит и эффекторная кодирующая способность патогенов Cochliobolus.PLoS Genet. https://doi.org/10.1371/journal.pgen.1003233
Артикул
PubMed
PubMed Central
Google Scholar
Cramer RA, Stajich JE, Yamanaka Y, Dietrich FS, Steinbach WJ, Perfect JR (2006) Филогеномный анализ нерибосомных пептидных синтетаз в роду Aspergillus . Gene 383: 24–32. https://doi.org/10.1016/j.gene.2006.07.008
CAS
Статья
PubMed
Google Scholar
Darriba D, Taboada GL, Doallo R, Posada D (2011) ProtTest 3: быстрый выбор наиболее подходящих моделей эволюции белка.Биоинформатика 27 (8): 1164–1165. https://doi.org/10.1093/bioinformatics/btr088
CAS
Статья
PubMed
PubMed Central
Google Scholar
de Hoog GS, Horre R (2002) Молекулярная таксономия видов Alternaria и Ulocladium от людей и их идентификация в обычной лаборатории. Микозы 45 (8): 259–276. https://doi.org/10.1046/j.1439-0507.2002.00747.Икс
Артикул
PubMed
Google Scholar
de Hoog GS, Zalar P, Gerrits van den Ende AHG, Gunde-Cimerman N (2005) Связь галотолерантности с патогенностью для человека в грибном древе жизни: обзор экологии и эволюции в условиях стресса. В: Гунде-Цимерман Н., Орен А., Племениташ А. (ред.) Адаптация к жизни при высоких концентрациях соли у архей, бактерий и эукарий. Springer, Dordrecht, pp 373–395
Google Scholar
de Hoog GS, Guarro J, Gené J, Figueras MJ (2015) Атлас клинических грибов.Лучший настольный инструмент для диагностики. https://doi.org/10.1111/j.1439-0507.1996.tb00148.x
de Hoog G, Ahmed S, Danesi P, Guillot J, Gräser Y (2018) Распространение патогенов и вспышек грибков в царстве грибов. In: Seyedmousavi S, de Hoog G, Guillot J, Verweij PE (eds) Новые и эпизоотические грибковые инфекции у животных. Springer, Dordrecht, pp 3–16
Google Scholar
de Jonge R, van Esse HP, Kombrink A, Shinya T, Desaki Y, Bours R, van der Krol S, Shibuya N, Joosten MHAJ, Thomma BPHJ (2010) Консервированный грибковый эффектор LysM Ecp6 предотвращает запускаемый хитином иммунитет в растениях.Наука 329 (5994): 953–955. https://doi.org/10.1126/science.11
CAS
Статья
PubMed
Google Scholar
de Waard MA, Andrade AC, Hayashi K, Schoonbeek HJ, Stergiopoulos I, Zwiers LH (2006) Влияние переносчиков грибковых лекарств на чувствительность к фунгицидам, множественную лекарственную устойчивость и вирулентность. Pest Manag Sci 62 (3): 195–207. https://doi.org/10.1002/ps.1150
CAS
Статья
PubMed
Google Scholar
Элиас PM (2007) Кожный барьер как элемент врожденного иммунитета.Семин Иммунопатол 29 (1): 3–14. https://doi.org/10.1007/s00281-007-0060-9
Артикул
PubMed
Google Scholar
Эллис М.Б. (1971) Демонические гифомицеты. CMI, Кью
Google Scholar
Feng B, Wang X, Hauser M, Kaufmann S, Jentsch S, Haase G, Becker JM, Szaniszlo PJ (2001) Молекулярное клонирование и характеристика WdPKS1 , гена, участвующего в дигидроксинафталине в биосинтезе меланина и Wangiella ( Exophiala ) dermatitidis .Infect Immun 69 (3): 1781–1794
CAS
Статья
Google Scholar
Фишбах М.А., Уолш СТ (2006) Сборочная энзимология поликетидных и нерибосомных пептидных антибиотиков: логика, механизмы и механизмы. Chem Rev 106 (8): 3468–3496. https://doi.org/10.1021/cr0503097
CAS
Статья
PubMed
Google Scholar
Galagan JE, Calvo SE, Borkovich KA, Selker EU, Read ND, Jaffe D, FitzHugh W, Ma LJ, Smirnov S, Purcell S, Rehman B, Elkins T, Engels R, Wang S, Nielsen CB, Butler J, Endrizzi M, Qui D, Ianakiev P, Bell-Pedersen D, Nelson MA, Werner-Washburne M, Selitrennikoff CP, Kinsey JA, Braun EL, Zelter A, Schulte U, Kothe GO, Jedd G, Mewes W, Staben C. , Marcotte E, Greenberg D, Roy A, Foley K, Naylor J, Stange-Thomann N, Barrett R, Gnerre S, Kamal M, Kamvysselis M, Mauceli E, Bielke C, Rudd S, Frishman D, Krystofova S, Rasmussen C , Метценберг Р.Л., Перкинс Д.Д., Крокен С., Когони С., Макино Дж., Кэтчесайд Д., Ли В., Пратт Р.Дж., Османи С.А., ДеСуза С.П., Гласс Л., Орбах М.Дж., Берглунд Дж.А., Фелькер Р., Ярден О., Пламанн М., Зайлер S, Dunlap J, Radford A, Aramayo R, Natvig DO, Alex LA, Mannhaupt G, Ebbole DJ, Freitag M, Paulsen I, Sachs MS, Lander ES, Nusbaum C, Birren B (2003) Последовательность генома нитчатого гриба Neurospora crassa .Nature 422 (6934): 859–868
CAS
Статья
Google Scholar
Гудвин С.Б., Бен М’Барек С., Диллон Б., Виттенберг А.Х., Крейн К.Ф., Хейн Дж. К., Фостер А.Дж., Ван дер Ли ТАД, Гримвуд Дж., Аэртс А., Антонив Дж., Бейли А., Блум Б., Боулер Дж. , Бристоу Дж., Ван дер Бургт А., Канто-Канче Б., Черчилль ACL, Конде-Ферраез Л., Коулс Х. Дж., Коутиньо П. М., Чукаи М., Дехал П., Де Вит П., Донцелли Б., Ван де Гест ХК, Ван Хэм РЧЖ, Хаммонд-Косак К.Е., Хенриссат Б., Килиан А., Кобаяши А.К., Купманн Е., Курмпетис Ю., Кузниар А., Линдквист Е., Ломбард В., Малипаард С., Мартинс Н., Мехраби Р., Нап JPH, Пономаренко А., Радд Дж. Schmutz J, Schouten HJ, Shapiro H, Stergiopoulos I, Torriani SFF, Tu H, de Vries RP, Waalwijk C, Ware SB, Wiebenga A, Zwiers LH, Оливер Р.П., Григорьев И.В., Кема GHJ (2011) Завершенный геном грибка Патоген пшеницы Mycosphaerella graminicola выявляет диспенсомную структуру, хромосомную пластичность и скрытый патогенез.PLoS Genet. https://doi.org/10.1371/journal.pgen.1002070
Артикул
PubMed
PubMed Central
Google Scholar
Gostinčar C, Grube M, de Hoog GS, Zalar P, Gunde-Cimerman N (2010) Экстремотолерантность у грибов: эволюция на грани. FEMS Microbiol Ecol 71 (1): 2–11
Артикул
Google Scholar
Gostinčar C, Grube M, Gunde-Cimerman N (2011) Эволюция грибковых патогенов в домашних условиях? Грибковая биол 115 (10): 1008–1018.https://doi.org/10.1016/j.funbio.2011.03.004
Артикул
PubMed
Google Scholar
Gostinčar C, Muggia L, Grube M (2012) Полиэкстремотолерантные черные грибы: олиготрофизм, адаптивный потенциал и связь с симбиозами лишайников. Фронтальный микробиол 3: 390. https://doi.org/10.3389/fmicb.2012.00390
Артикул
PubMed
PubMed Central
Google Scholar
Gostinčar C, Ohm RA, Kogej T, Sonjak S, Turk M, Zajc J, Zalar P, Grube M, Sun H, Han J, Sharma A, Chiniquy J, Ngan CY, Lipzen A, Barry K, Григорьев IV, Gunde-Cimerman N (2014) Секвенирование генома четырех сортов Aureobasidium pullulans : биотехнологический потенциал, стрессоустойчивость и описание новых видов.BMC Genomics 15 (1): 549. https://doi.org/10.1186/1471-2164-15-549
CAS
Статья
PubMed
PubMed Central
Google Scholar
Gostinčar C, Gunde-Cimerman N, Grube M (2015) Экстремотолерантность как ответ грибов на изменение окружающей среды. В: Bakermans C (ed) Микробная эволюция в экстремальных условиях. Жизнь в экстремальных условиях, т. 2. de Gruyter, Berlin, pp. 185–208. https: // doi.org / 10.1515 / 9783110340716.185
Григорьев И.В., Никитин Р., Харидас С., Куо А., Ом Р., Отиллар Р., Райли Р., Саламов А., Жао Х, Корзеневский Ф., Смирнова Т., Нордберг Н., Дубчак И., Шабалов И. (2014) Портал MycoCosm : готовимся к 1000 геномов грибов. Nucleic Acids Res 42 (1): 699–704. https://doi.org/10.1093/nar/gkt1183
CAS
Статья
Google Scholar
Gugnani HC, Okafor JI (1980) Микотическая флора кишечника и других внутренних органов некоторых рептилий и земноводных с особым упором на характеристику изолятов Basidiobolus.Mykosen 23 (5): 260–268
CAS
Статья
Google Scholar
Guillot J, Demanche C, Hugot JP, Berthelemy M, Wakefield E, Dei-Cas E, Chermette R (2001) Параллельные филогении видов Pneumocystis и их млекопитающих-хозяев. J Eukaryot Microbiol 48 (1): 113–115
Статья
Google Scholar
Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W., Gascuel O (2010) Новые алгоритмы и методы для оценки филогении максимального правдоподобия: оценка производительности PhyML 3.0. Syst Biol 59 (3): 307–321. https://doi.org/10.1093/sysbio/syq010
CAS
Статья
PubMed
Google Scholar
Haese A, Schubert M, Herrmann M, Zocher R (1993) Молекулярная характеристика гена энниатин синтетазы, кодирующего многофункциональный фермент, катализирующий образование N-метилдепсипептида в Fusarium scirpi . Mol Microbiol 7 (6): 905–914. https://doi.org/10.1111/j.1365-2958.1993.tb01181.x
CAS
Статья
PubMed
Google Scholar
Hall TA (1999) BioEdit: удобный редактор выравнивания биологических последовательностей и программа анализа для Windows 95/98 / NT. Нуклеиновые кислоты Symp Ser 41: 95–98
CAS
Google Scholar
Hamad M (2008) Противогрибковая иммунотерапия и иммуномодуляция: двойной подход к борьбе с инвазивными грибковыми инфекциями.Scand J Immunol 67 (6): 533–543. https://doi.org/10.1111/j.1365-3083.2008.02101.x
CAS
Статья
PubMed
Google Scholar
Hamada N, Abe N (2010) Сравнение грибков, обнаруженных в ванных комнатах и раковинах. Biocontrol Sci 15 (2): 51–56
Статья
Google Scholar
Han MV, Thomas GWC, Lugo-Martinez J, Hahn MW (2013) Оценка скорости роста и потери генов при наличии ошибки сборки и аннотации генома с использованием CAFE 3.Mol Biol Evol 30 (8): 1987–1997. https://doi.org/10.1093/molbev/mst100
CAS
Статья
PubMed
Google Scholar
Jung WH, Sham A, White R, Kronstad JW (2006) Железная регуляция основных факторов вирулентности у СПИД-ассоциированного патогена Cryptococcus neoformans . PLoS Biol 4 (12): 2282–2295. https://doi.org/10.1371/journal.pbio.0040410
CAS
Статья
Google Scholar
Karanyi Z, Holb I, Hornok L, Pocsi I, Miskei M (2013) FSRD: база данных грибковой реакции на стресс.База данных. https://doi.org/10.1093/database/bat037
Артикул
PubMed
PubMed Central
Google Scholar
Katoh K, Toh H (2008) Последние разработки в программе множественного выравнивания последовательностей MAFFT. Краткий биоинформ 9 (4): 286–298
CAS
Статья
Google Scholar
Килинг КБ, Павур Р.Дж. (2011) Статистическая точность программного обеспечения для работы с электронными таблицами.Am Stat 65 (4): 265–273. https://doi.org/10.1198/tas.2011.09076
Артикул
Google Scholar
Кейжар А., Гобец С., Племениташ А., Ленасси М. (2013) Меланин имеет решающее значение для роста черных дрожжей Hortaea werneckii в естественной гиперсоленой среде. Fungal Biol 117 (5): 368–379. https://doi.org/10.1016/j.funbio.2013.03.006
CAS
Статья
PubMed
Google Scholar
Kempf VAJ, Hitziger N, Riess T, Autenrieth IB (2002) Имеют ли патогены растений и человека общую стратегию патогенности? Trends Microbiol 10 (6): 269–275.https://doi.org/10.1016/S0966-842x(02)02372-7
CAS
Статья
PubMed
Google Scholar
Kirzinger MWB, Nadarasah G, Stavrinides J (2011) Понимание патогенных бактерий, вызывающих различные виды растений. Гены 2 (4): 980–997. https://doi.org/10.3390/genes2040980
Артикул
PubMed
PubMed Central
Google Scholar
Кис-Папо Т., Вейг А.Р., Райли Р., Персох Д., Саламов А., Сан Х., Липзен А., Вассер С.П., Рамбольд Г., Григорьев И.В., Нево Е. (2014) Геномные адаптации галофильных нитчатых грибов Мертвого моря Eurotium rubrum .Nat Commun. https://doi.org/10.1038/ncomms4745
Артикул
PubMed
Google Scholar
Когей Т., Штейн М., Фолькманн М., Горбушина А.А., Галински Е.А., Гунде-Цимерман Н. (2007) Осмотическая адаптация галофильного гриба Hortaea werneckii : роль осмолитов и меланизация. Microbiology 153 (Pt 12): 4261–4273
CAS
Статья
Google Scholar
Krijger JJ, Thon MR, Deising HB, Wirsel SGR (2014) Состав грибковых секретов указывает на большее влияние филогенетического анамнеза, чем адаптация к образу жизни.BMC Genomics. https://doi.org/10.1186/1471-2164-15-722
Артикул
PubMed
PubMed Central
Google Scholar
Krogh A, Larsson B, von Heijne G, Sonnhammer ELL (2001) Прогнозирование топологии трансмембранного белка с помощью скрытой марковской модели: применение для полных геномов. J Mol Biol 305 (3): 567–580. https://doi.org/10.1006/jmbi.2000.4315
CAS
Статья
PubMed
Google Scholar
Kumamoto CA (2008) Экспрессия гена, специфичная для ниши в течение C.albicans инфекции. Curr Opin Microbiol 11 (4): 325–330. https://doi.org/10.1016/j.mib.2008.05.008
CAS
Статья
PubMed
PubMed Central
Google Scholar
Langfelder K, Streibel M, Jahn B, Haase G, Brakhage AA (2003) Биосинтез меланинов грибов и их значение для патогенных грибов человека. Fungal Genet Biol 38 (2): 143–158. https://doi.org/10.1016/S1087-1845(02)00526-1
CAS
Статья
PubMed
Google Scholar
Lenassi M, Gostinčar C, Jackman S, Turk M, Sadowski I, Nislow C, Jones S, Birol I, Gunde-Cimerman N, Plemenitaš A (2013) Дупликация всего генома и обогащение переносчиков катионов металлов выявлено de novo секвенирование генома чрезвычайно галотолерантных черных дрожжей Hortaea werneckii .PLoS ONE 8 (8): e71328. https://doi.org/10.1371/journal.pone.0071328
CAS
Статья
PubMed
PubMed Central
Google Scholar
Lian X, de Hoog GS (2010) Влажные клетки в помещении содержат меланизированные агенты кожной инфекции. Med Mycol 48 (4): 622–628. https://doi.org/10.3109/13693780
5774
CAS
Статья
PubMed
Google Scholar
Liu L, Zhang Z, Shao CL, Wang JL, Bai H, Wang CY (2015) Биоинформатический анализ последовательностей, структур и функций матричных доменов грибных продуктов поликетидсинтазы.Научный представитель https://doi.org/10.1038/srep10463
Артикул
PubMed
PubMed Central
Google Scholar
Lowe RGT, McCorkelle O, Bleackley M, Collins C, Faou P, Mathivanan S, Anderson M (2015) Внеклеточные пептидазы злокачественного патогена Fusarium graminearum . Фронтальный завод им. https://doi.org/10.3389/fpls.2015.00962
Артикул
PubMed
PubMed Central
Google Scholar
Lu T, Yao B, Zhang C (2012) DFVF: база данных факторов вирулентности грибов.База данных. https://doi.org/10.1093/database/bas032
Артикул
PubMed
PubMed Central
Google Scholar
Melotto M, Panchal S, Roy D (2014) Врожденный иммунитет растений против бактериальных патогенов человека. Front Microbiol. https://doi.org/10.3389/fmicb.2014.00411
Артикул
PubMed
PubMed Central
Google Scholar
Миллер М., Пфайфер В., Шварц Т. (2010) Создание научного портала CIPRES для вывода больших филогенетических деревьев.В: Proceedings of the Gateway Computing Environments Workshop (GCE), vol 1 (1), pp 1-8
Monod M, Capoccia S, Lechenne B, Zaugg C, Holdom M, Jousson O (2002) Секретные протеазы из патогенные грибы. Int J Med Microbiol 292 (5–6): 405–419. https://doi.org/10.1078/1438-4221-00223
CAS
Статья
PubMed
Google Scholar
Носанчук JD, Stark RE, Casadevall A (2015) Грибковый меланин: что мы знаем о структуре? Front Microbiol.https://doi.org/10.3389/fmicb.2015.01463
Артикул
PubMed
PubMed Central
Google Scholar
Новак Бабич М., Залар П., Женко Б., Джероски С., Гунде-Цимерман Н. (2016) Дрожжи и дрожжеподобные грибы в водопроводной и грунтовой воде и их передача бытовым приборам. Грибковые экологии 20: 30–39. https://doi.org/10.1016/j.funeco.2015.10.001
Артикул
Google Scholar
O’Hanlon KA, Gallagher L, Schrettl M, Jochl C, Kavanagh K, Larsen TO, Doyle S (2012) Нерибосомные гены пептид синтетазы pesL и pes1 необходимы для производства фумигаклавина C в Aspergillus Aspergillus fumigatus.Appl Environ Microbiol 78 (9): 3166–3176. https://doi.org/10.1128/Aem.07249-11
Артикул
PubMed
PubMed Central
Google Scholar
Ом RA, Feau N, Henrissat B, Schoch CL, Horwitz BA, Barry KW, Condon BJ, Copeland AC, Dhillon B, Glaser F, Hesse CN, Kosti I, Labutti K, Lindquist EA, Lucas S, Salamov AA, Bradshaw RE, Ciuffetti L, Hamelin RC, Kema GH, Lawrence C, Scott JA, Spatafora JW, Turgeon BG, de Wit PJ, Zhong S, Goodwin SB, Григорьев И.В. (2012) Разнообразный образ жизни и стратегии патогенеза растений, закодированные в геномы восемнадцати грибов Dothideomycetes.PLoS Pathog 8 (12): e1003037. https://doi.org/10.1371/journal.ppat.1003037
CAS
Статья
PubMed
PubMed Central
Google Scholar
Oren A (2011) Термодинамические пределы микробной жизни при высоких концентрациях соли. Environ Microbiol 13 (8): 1908–1923. https://doi.org/10.1111/j.1462-2920.2010.02365.x
CAS
Статья
PubMed
Google Scholar
Park BH, Karpinets TV, Syed MH, Leuze MR, Uberbacher EC (2010) CAZymes Analysis Toolkit (CAT): веб-сервис для поиска и анализа углеводно-активных ферментов в недавно секвенированном организме с использованием базы данных CAZy.Гликобиология 20 (12): 1574–1584. https://doi.org/10.1093/glycob/cwq106
CAS
Статья
PubMed
Google Scholar
Pel HJ, de Winde JH, Archer DB, Dyer PS, Hofmann G, Schaap PJ, Turner G, de Vries RP, Albang R, Albermann K, Andersen MR, Bendtsen JD, Benen JA, van den Berg M, Breestraat S, Caddick MX, Contreras R, Cornell M, Coutinho PM, Danchin EG, Debets AJ, Dekker P, van Dijck PW, van Dijk A, Dijkhuizen L, Driessen AJ, d’Enfert C, Geysens S, Goosen C, Groot GS, de Groot PW, Guillemette T., Henrissat B, Herweijer M, van den Hombergh JP, van den Hondel CA, van der Heijden RT, van der Kaaij RM, Klis FM, Kools HJ, Kubicek CP, van Kuyk PA, Lauber J , Лу Х, ван дер Маарель М.Дж., Меуленберг Р., Менке Х., Мортимер М.А., Нильсен Дж., Оливер С.Г., Олстхорн М., Пал К., ван Пей Н.Н., Рам А.Ф., Ринас Ю.А., Рубос Д.А., Сагт К.М., Шмоль М., Вс J, Ussery D, Varga J, Vervecken W, van de Vondervoort PJ, Wedler H, Wosten HA, Zeng AP, van Ooyen AJ, Visser J, Stam H (2007) Секвенирование генома и анализ универсальной фабрики клеток Aspergillus niger КОС 513.88. Nat Biotechnol 25 (2): 221–231
Статья
Google Scholar
Petersen TN, Brunak S, von Heijne G, Nielsen H (2011) SignalP 4.0: различение сигнальных пептидов из трансмембранных областей. Нат Методы 8 (10): 785–786. https://doi.org/10.1038/nmeth.1701
CAS
Статья
PubMed
Google Scholar
Philpott CC (2006) Поглощение железа грибами: система для каждого источника.BBA Mol Cell Res 1763 (7): 636–645. https://doi.org/10.1016/j.bbamcr.2006.05.008
CAS
Статья
Google Scholar
Prenafeta-Boldu FX, Summerbell R, de Hoog GS (2006) Грибы, растущие на ароматических углеводородах: неожиданная встреча биотехнологии с биологической опасностью? FEMS Microbiol Rev 30 (1): 109–130. https://doi.org/10.1111/j.1574-6976.2005.00007.x
CAS
Статья
PubMed
Google Scholar
Prenafeta-Boldu FX, Guivernau M, Gallastegui G, Vinas M, de Hoog GS, Elias A (2012) Грибковые / бактериальные взаимодействия во время биоразложения углеводородов TEX (толуол, этилбензол и п-ксилол) в работающих газовых биофильтрах в ксерофильных условиях.FEMS Microbiol Ecol 80 (3): 722–734. https://doi.org/10.1111/j.1574-6941.2012.01344.x
CAS
Статья
PubMed
Google Scholar
Punta M, Coggill PC, Eberhardt RY, Mistry J, Tate J, Boursnell C, Pang N, Forslund K, Ceric G, Clements J, Heger A, Holm L, Sonnhammer ELL, Eddy SR, Bateman A, Finn RD (2012) База данных семейств белков Pfam. Nucleic Acids Res 40: 290–301. https://doi.org/10.1093/Nar/Gkr1065
Артикул
Google Scholar
R Development Core Team (2015) R: язык и среда для статистических вычислений.Фонд R для статистических вычислений, Вена
Google Scholar
Ranganathan S, Garg G (2009) Секретом: ключи к патогенной инфекции и клиническому применению. Genome Med. https://doi.org/10.1186/gm113
Артикул
PubMed
PubMed Central
Google Scholar
Rawlings ND, Barrett AJ, Bateman A (2012) MEROPS: база данных протеолитических ферментов, их субстратов и ингибиторов.Nucleic Acids Res 40 (D1): 343–350. https://doi.org/10.1093/Nar/Gkr987
Артикул
Google Scholar
Reichard U, Lechenne B, Asif AR, Streit F, Grouzmann E, Jousson O, Monod M (2006) Седолизины, новый класс секретируемых протеаз из aspergillus fumigatus с эндопротеазной или трипептидилпептидазной активностью при кислых pH. Appl Environ Microbiol 72 (3): 1739–1748. https://doi.org/10.1128/Aem.72.3.1739-1748.2006 г.
CAS
Статья
PubMed
PubMed Central
Google Scholar
Robbertse B, Yoder RJ, Boyd A, Reeves J, Spatafora JW (2011) Hal: автоматизированный конвейер для филогенетического анализа геномных данных. PLoS Curr 3: RRN213. https://doi.org/10.1371/currents.rrn1213
Артикул
Google Scholar
Роберт В.А., Casadevall A (2009) Эндотермия позвоночных ограничивает большинство грибов как потенциальных патогенов.J Infect Dis 200 (10): 1623–1626. https://doi.org/10.1086/644642
Артикул
PubMed
Google Scholar
Sanchez-Vallet A, Saleem-Batcha R, Kombrink A, Hansen G, Valkenburg DJ, Thomma BPHJ, Mesters JR (2013) Грибковый эффектор Ecp6 превосходит иммунный рецептор хозяина для связывания хитина посредством внутрицепной димеризации LysM. Элиф. https://doi.org/10.7554/elife.00790
Артикул
PubMed
PubMed Central
Google Scholar
Sanderson MJ (2003) r8s: определение абсолютных скоростей молекулярной эволюции и времени расхождения в отсутствие молекулярных часов.Биоинформатика 19 (2): 301–302. https://doi.org/10.1093/bioinformatics/19.2.301
CAS
Статья
PubMed
Google Scholar
Schmaler-Ripcke J, Sugareva V, Gebhardt P, Winkler R, Kniemeyer O, Heinekamp T, Brakhage AA (2009) Производство пиомеланина, второго типа меланина, через путь деградации тирозина в Aspergillus fumigatus Aspergillus fumigatus . Appl Environ Microbiol 75 (2): 493–503. https: // doi.org / 10.1128 / Aem.02077-08
CAS
Статья
PubMed
Google Scholar
Schnitzler N, Peltroche-Llacsahuanga H, Bestier N, Zundorf J, Lutticken R, Haase G (1999) Влияние меланина и каротиноидов из Exophiala ( Wangiella ) на дерматитизоцит, окислительно-восстановительный убийство нейтрофилами человека. Инфекция иммунной 67 (1): 94–101
CAS
PubMed
PubMed Central
Google Scholar
Schoch CL, Crous PW, Groenewald JZ, Boehm EWA, Burgess TI, de Gruyter J, de Hoog GS, Dixon LJ, Grube M, Gueidan C, Harada Y, Hatakeyama S, Hirayama K, Hosoya SM T, Huhndorf , Hyde KD, Jones EBG, Kohlmeyer J, Kruys A, Li YM, Lucking R, Lumbsch HT, Marvanova L, Mbatchou JS, Mcvay AH, Miller AN, Mugambi GK, Muggia L, Nelsen MP, Nelson P, Owensby CA, Phillips AJL, Phongpaichit S, Pointing SB, Pujade-Renaud V, Raja HA, Plata ER, Robbertse B, Ruibal C, Sakayaroj J, Sano T, Selbmann L, Shearer CA, Shirouzu T, Slippers B, Suetrong S, Tanaka K, Volkmann -Kohlmeyer B, Wingfield MJ, Wood AR, Woudenberg JHC, Yonezawa H, Zhang Y, Spatafora JW (2009) Филогенетическая оценка дотидеомицетов в масштабах всего класса.Стад Микол 64: 1–15. https://doi.org/10.3114/sim.2009.64.01
CAS
Статья
PubMed
PubMed Central
Google Scholar
Segorbe D, Di Pietro A, Perez-Nadales E, Turra D (2017) Три митоген-активируемые протеинкиназы (MAPK) Fusarium oxysporum играют различные и взаимодополняющие роли в адаптации к стрессу и патогенности между царствами. Mol Plant Pathol 18 (7): 912–924. https: // doi.org / 10.1111 / mpp.12446
CAS
Статья
PubMed
Google Scholar
Sharma K, Goss EM, Dickstein ER, Smith ME, Johnson JA, Southwick FS, van Bruggen AHC (2014) Exserohilum rostratum : характеристика патогена растений и человека между разными царствами. PLoS ONE. https://doi.org/10.1371/journal.pone.0108691
Артикул
PubMed
PubMed Central
Google Scholar
Silva MG, Schrank A, Bailao EFLC, Bailao AM, Borges CL, Staats CC, Parente JA, Pereira M, Salem-Izacc SM, Mendes-Giannini MJS, Oliveira RMZ, Silva LKRE, Nosanchuk JD, Vainstein MH Soares CMD (2011) Гомеостаз железа, меди и цинка в Paracoccidioides brasiliensis , Cryptococcus neoformans var. grubii и Cryptococcus gattii : сравнительный анализ. Front Microbiol. https://doi.org/10.3389/fmicb.2011.00049
Артикул
PubMed
PubMed Central
Google Scholar
Silveira F, Nucci M (2001) Появление черной плесени при грибковых заболеваниях: эпидемиология и терапия. Curr Opin Infect Dis 14 (6): 679–684
CAS
Статья
Google Scholar
Singh A, Rella A, Schwacke J, Vacchi-Suzzi C, Luberto C, Del Poeta M (2015) Экспрессия трансмембранного переносчика, регулируемая глюкозилцерамидным путем в Cryptococcus neoformans .Примечания BMC Res 8 (1): 681. https://doi.org/10.1186/s13104-015-1613-y
CAS
Статья
PubMed
PubMed Central
Google Scholar
Sinha S, Flibotte S, Neira M, Formby S, Plemenitas A, Cimerman NG, Lenassi M, Gostincar C, Stajich JE, Nislow C (2017) Анализ недавней дупликации генома галофильных дрожжей Hortaea 1664 werneckii : сочетание улучшенного генома с экспрессией генов и структурой хроматина.G3 Genes Genom Genet 7 (7): 2015–2022. https://doi.org/10.1534/g3.117.040691
Артикул
Google Scholar
Šišić A, Baćanović-Šišić J, Al-Hatmi AMS, Karlovsky P, Ahmed SA, Maier W., de Hoog GS, Finckh MR (2018) Выпуск ‘forma specialis’ в Fusarium : тематическое исследование в Fusarium solani f. sp. писи . Научный представитель https://doi.org/10.1038/s41598-018-19779-z
Артикул
PubMed
PubMed Central
Google Scholar
Слепецки Р.А., Стармер В.Т. (2009) Фенотипическая пластичность грибов: обзор наблюдений за Aureobasidium pullulans .Микология 101 (6): 823–832. https://doi.org/10.3852/08-197
Артикул
PubMed
Google Scholar
Song YG, Laureijssen-van de Sande WWJ, Moreno LF, van den Ende BG, Li RY, de Hoog S (2017) Сравнительная экология капсульных видов Exophiala , вызывающих диссеминированную инфекцию у людей. Front Microbiol. https://doi.org/10.3389/fmicb.2017.02514
Артикул
PubMed
PubMed Central
Google Scholar
Talavera G, Castresana J (2007) Улучшение филогении после удаления расходящихся и неоднозначно выровненных блоков из выравнивания последовательностей белков.Syst Biol 56 (4): 564–577. https://doi.org/10.1080/10635150701472164
CAS
Статья
PubMed
Google Scholar
ten Have A, Dekkers E, Kay J, Phylip LH, van Kan JAL (2004) Семейство генов аспарагиновой протеиназы в мицелиальных грибах Botrytis cinerea содержит членов с новыми характеристиками. Микробиология 150: 2475–2489. https://doi.org/10.1099/mic.0.27058-0
CAS
Статья
PubMed
Google Scholar
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) Интерфейс окон ClustalX: гибкие стратегии для множественного выравнивания последовательностей с помощью инструментов анализа качества.Nucleic Acid Res 24: 4876–4882
Статья
Google Scholar
van Baarlen P, van Belkum A, Summerbell RC, Crous PW, Thomma BPHJ (2007) Молекулярные механизмы патогенности: как патогенные микроорганизмы развивают скачки хозяина между царствами? FEMS Microbiol Rev 31 (3): 239–277. https://doi.org/10.1111/j.1574-6976.2007.0065.x
Артикул
PubMed
Google Scholar
van Burik JAH, Magee PT (2001) Аспекты грибкового патогенеза у людей.Annu Rev Microbiol 55: 743–772
Статья
Google Scholar
Varga J, Rigo K, Kocsube S, Farkas B, Pal K (2003) Разнообразие последовательностей генов поликетидсинтазы у видов Aspergillus . Res Microbiol 154 (8): 593–600. https://doi.org/10.1016/S0923-2508(03)00169-4
CAS
Статья
PubMed
Google Scholar
von Döhren H (2009) Обзор генов нерибосомальной пептидной синтетазы (NRPS) в Aspergillus nidulans .Fungal Genet Biol 46: S45 – S52. https://doi.org/10.1016/j.fgb.2008.08.008
CAS
Статья
Google Scholar
Walton JD (1996) Хоз-селективные токсины: агенты совместимости. Растительная клетка 8 (10): 1723–1733
CAS
Статья
Google Scholar
Залар П., Новак М., Де Хуг Г.С., Гунде-Цимерман Н. (2011) Посудомоечные машины — искусственная экологическая ниша, вмещающая условно-патогенные грибковые патогены человека.Грибковая биол 115 (10): 997–1007. https://doi.org/10.1016/j.funbio.2011.04.007
CAS
Статья
PubMed
Google Scholar
Ziemert N, Podell S, Penn K, Badger JH, Allen E, Jensen PR (2012) Искатель области естественного продукта NaPDoS: основанный на филогенезе биоинформатический инструмент для классификации разнообразия генов вторичных метаболитов. PLoS ONE. https://doi.org/10.1371/journal.pone.0034064
Артикул
PubMed
PubMed Central
Google Scholar
Зупанчич Ю., Бабич М.Н., Залар П., Гунде-Цимерман Н. (2016) Черные дрожжи Exophiala dermatitidis и другие избранные условно-патогенные грибковые патогены человека распространяются из посудомоечных машин на кухню.PLoS ONE. https://doi.org/10.1371/journal.pone.0148166
Артикул
PubMed
PubMed Central
Google Scholar
Последовательность генома насекомого-патогенного гриба Cordyceps militaris, ценного объекта традиционной китайской медицины | Геномная биология
Сунг Г.Х., Хиуэл-Джонс Н.Л., Сунг Дж. М., Луангса-Ард Дж. Дж., Шреста Б., Спатафора Дж. У .: Филогенетическая классификация кордицепса и ключичных грибов.Stud Mycol. 2007, 57: 5-59. 10.3114 / sim.2007.57.01.
PubMed
PubMed Central
Статья
Google Scholar
de Faria MR, Wraight SP: Mycoinsecticides и Mycoacaricides: исчерпывающий список с мировым охватом и международной классификацией типов рецептур. Биол Контроль. 2007, 43: 237-256. 10.1016 / j.biocontrol.2007.08.001.
CAS
Статья
Google Scholar
St Leger RJ, Wang C: Генная инженерия агентов биоконтроля грибков для достижения большей эффективности против насекомых-вредителей. Appl Microbiol Biotechnol. 2010, 85: 901-907. 10.1007 / s00253-009-2306-z.
PubMed
CAS
Статья
Google Scholar
Патерсон RR: Кордицепс : традиционная китайская медицина и еще одна биофабрика по лечению грибков. Фитохимия. 2008, 69: 1469-1495. 10.1016 / j.phytochem.2008.01.027.
PubMed
CAS
Статья
Google Scholar
Чжоу Х, Гун З, Су И, Лин Дж, Тан К: Кордицепс грибы: натуральные продукты, фармакологические функции и продукты для развития. J Pharm Pharmacol. 2009, 61: 279-291.
PubMed
CAS
Статья
Google Scholar
Сяо Дж. Х., Чжун Дж. Дж.: Вторичные метаболиты из видов кордицепса и исследования их противоопухолевой активности.Последние публикации Pat Biotechnol. 2007, 1: 123-137. 10.2174 / 187220807780809454.
PubMed
CAS
Статья
Google Scholar
ClinicalTrials.gov. [http://clinicaltrials.gov/show/NCT00709215]
Stone R: Последний бой похитителя тел в Гималаях ?. Наука. 2008, 322: 1182-10.1126 / science.322.5905.1182.
PubMed
CAS
Статья
Google Scholar
Ли SC, Ni M, Li W, Shertz C, Heitman J: Эволюция пола: взгляд из царства грибов. Microbiol Mol Biol Rev.2010, 74: 298-340. 10.1128 / MMBR.00005-10.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Galagan JE, Calvo SE, Cuomo C, Ma LJ, Wortman JR, Batzoglou S, Lee SI, Bastürkmen M, Spevak CC, Clutterbuck J, Kapitonov V, Jurka J, Scazzocchio C, Farman M, Butler J , Перселл С., Харрис С., Браус Г. Х., Драт О, Буш С., Д’Энферт С., Бушье С., Голдман Г. Х., Белл-Педерсен Д., Гриффитс-Джонс С., Дунан Дж. Х., Ю Дж., Виенкен К., Пейн А., Фрайтаг M, et al: Секвенирование Aspergillus nidulans и сравнительный анализ с A.fumigatus и A. oryzae . Природа. 2005, 438: 1105-1115. 10.1038 / природа04341.
PubMed
CAS
Статья
Google Scholar
Гао Q, Цзинь К., Инь Ш., Чжан И, Сяо Г, Шан Y, Дуань З., Ху X, Се XQ, Чжоу Г, Пэн Г, Ло З, Хуанг В., Ван Б, Фанг В. , Wang S, Zhong Y, Ma LJ, St Leger RJ, Zhao GP, Pei Y, Feng MG, Xia Y, Wang CS: Секвенирование генома и сравнительная транскриптомика модельных энтомопатогенных грибов Metarhizium anisopliae и M.акридум . PLoS Genet. 2011, 7: e1001264-10.1371 / journal.pgen.1001264.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Xiong CH, Xia YL, Zheng P, Shi SH, Wang CS: Профилирование экспрессии генов, специфичных для стадии развития, для лекарственного гриба Cordyceps militaris . Микология. 2010, 1: 25-66.
CAS
Статья
Google Scholar
Galagan JE, Calvo SE, Borkovich KA, Selker EU, Read ND, Jaffe D, FitzHugh W, Ma LJ, Smirnov S, Purcell S, Rehman B, Elkins T, Engels R, Wang S, Nielsen CB, Butler J, Endrizzi M, Qui D, Ianakiev P, Bell-Pedersen D, Nelson MA, Werner-Washburne M, Selitrennikoff CP, Kinsey JA, Braun EL, Zelter A, Schulte U, Kothe GO, Jedd G, Mewes W и др.: Геном последовательность нитчатого гриба Neurospora crassa . Природа. 2003, 422: 859-868. 10.1038 / природа01554.
PubMed
CAS
Статья
Google Scholar
Ma LJ, van der Does HC, Borkovich KA, Coleman JJ, Daboussi MJ, Di Pietro A, Dufresne M, Freitag M, Grabherr M, Henrissat B, Houterman PM, Kang S, Shim WB, Woloshuk C, Xie X, Xu JR, Antoniw J, Baker SE, Bluhm BH, Breakspear A, Brown DW, Butchko RA, Chapman S, Coulson R, Coutinho PM, Danchin EG, Diener A, Gale LR, Gardiner DM, Goff S и др.: Сравнительная геномика показывает подвижные хромосомы патогенности в Fusarium . Природа. 2010, 464: 367-373. 10.1038 / природа08850.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Kubicek CP, Herrera-Estrella A, Seidl-Seiboth V, Martinez DA, Druzhinina IS, Thon M, Zeilinger S, Casas-Flores S, Horwitz BA, Mukherjee PK, Mukherjee M, Kredics L, Alcaraz LD, Aerts A, Antal З., Атанасова Л., Сервантес-Бадильо М.Г., Чалакомб Дж., Чертков О., МакКласки К., Кулпье Ф., Дешпанде Н., фон Дёрен Х., Эбболе DJ, Эскивель-Наранхо ЕС, Фекете Э, Флиппи М., Глейзер Ф, Гомес-Родригес , Gruber S и др.: Сравнительный анализ последовательности генома подчеркивает микопаразитизм как унаследованный от предков образ жизни Trichoderma .Genome Biol. 2011, 12: R40-10.1186 / gb-2011-12-4-r40.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Раджашекара Дж., Гласнер Дж. Д., Гловер Д.А., Сплиттер Г.А.: Сравнительная полногеномная гибридизация выявила геномные островки у видов Brucella . J Bacteriol. 2004, 186: 5040-5051. 10.1128 / JB.186.15.5040-5051.2004.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Fedorova ND, Khaldi N, Joardar VS, Maiti R, Amedeo P, Anderson MJ, Crabtree J, Silva JC, Badger JH, Albarraq A, Angiuoli S, Bussey H, Bowyer P, Cotty PJ, Dyer PS, Egan A, Galens K, Fraser-Liggett CM, Haas BJ, Inman JM, Kent R, Lemieux S, Malavazi I, Orvis J, Roemer T., Ronning CM, Sundaram JP, Sutton G, Turner G, Venter JC и др.: Геномные острова в Патогенный мицелиальный гриб Aspergillus fumigatus . PLoS Genet. 2008, 4: e1000046-10.1371 / journal.pgen.1000046.
PubMed
PubMed Central
Статья
Google Scholar
McElwain JC, Punyasena SW: События массового вымирания и летопись окаменелостей растений. Trends Ecol Evol. 2007, 22: 548-557. 10.1016 / j.tree.2007.09.003.
PubMed
Статья
Google Scholar
Cuomo CA, Güldener U, Xu JR, Trail F, Turgeon BG, Di Pietro A, Walton JD, Ma LJ, Baker SE, Rep M, Adam G, Antoniw J, Baldwin T, Calvo S, Chang YL, Decaprio D, Gale LR, Gnerre S, Goswami RS, Hammond-Kosack K, Harris LJ, Hilburn K, Kennell JC, Kroken S, Magnuson JK, Mannhaupt G, Mauceli E, Mewes HW, Mitterbauer R, Muehlbauer G и др. al: Геном Fusarium graminearum показывает связь между локализованным полиморфизмом и специализацией патогенов.Наука. 2007, 317: 1400-1402. 10.1126 / science.1143708.
PubMed
CAS
Статья
Google Scholar
Галаган Дж. Э., Селкер ЕС: RIP: эволюционная цена защиты генома. Тенденции Genet. 2004, 20: 417-423. 10.1016 / j.tig.2004.07.007.
PubMed
CAS
Статья
Google Scholar
Screen SE, St Leger RJ: Клонирование, экспрессия и субстратная специфичность грибкового химотрипсина.Доказательства латерального переноса гена от бактерии-актиномицета. J Biol Chem. 2000, 275: 6689-6694. 10.1074 / jbc.275.9.6689.
PubMed
CAS
Статья
Google Scholar
Coetzer TH, Goldring JP, Huson LE: Олигопептидаза B: процессинговая пептидаза, участвующая в патогенезе. Биохимия. 2008, 90: 336-344. 10.1016 / j.biochi.2007.10.011.
PubMed
CAS
Статья
Google Scholar
Chong SL, Battaglia E, Coutinho PM, Henrissat B, Tenkanen M, de Vries RP: α-глюкуронидаза Agu1 из Schizophyllum commune является членом нового семейства гликозидгидролаз (Gh215). Appl Microbiol Biotechnol. 2011, 90: 1323-1332. 10.1007 / s00253-011-3157-у.
PubMed
CAS
Статья
Google Scholar
Дуань З.Б., Шан И, Гао К., Чжэн П., Ван К.С.: Фосфокетолаза Mpk1 бактериального происхождения адаптивно необходима для полной вирулентности в патогенном грибке, вызывающем насекомых Metarhizium anisopliae .Environ Microbiol. 2009, 11: 2351-2360. 10.1111 / j.1462-2920.2009.01961.x.
PubMed
CAS
Статья
Google Scholar
Crešnar B, Petrič S: Ферменты цитохрома P450 в царстве грибов. Biochim Biophys Acta. 2011, 1814: 29-35.
PubMed
Статья
Google Scholar
Келлер Н.П., Тернер Дж., Беннетт Дж. В.: Вторичный метаболизм грибов — от биохимии к геномике.Nat Rev Microbiol. 2005, 3: 937-947. 10.1038 / nrmicro1286.
PubMed
CAS
Статья
Google Scholar
Джарролд С.Л., Мур Д., Поттер У., Чарнли А.К .: Вклад поверхностных восков в рост энтомопатогенных грибов до проникновения на кутикулу хозяина. Mycol Res. 2007, 111: 240-249. 10.1016 / j.mycres.2006.10.007.
PubMed
Статья
Google Scholar
Morschhauser J: Регулирование множественной лекарственной устойчивости патогенных грибов. Fungal Genet Biol. 2010, 47: 94-106. 10.1016 / j.fgb.2009.08.002.
PubMed
Статья
Google Scholar
Luz C, Netto MC, Rocha LF: Чувствительность in vitro к фунгицидам, вызываемым беспозвоночными патогенными и сапробными грибами. Микопатология. 2007, 164: 39-47. 10.1007 / s11046-007-9020-0.
PubMed
CAS
Статья
Google Scholar
Kulkarni RD, Thon MR, Pan H, Dean RA: Новые рецепторы-подобные белки, связанные с G-белками, в патогенном грибке растений Magnaporthe grisea . Genome Biol. 2005, 6: R24-10.1186 / GB-2005-6-3-r24.
PubMed
PubMed Central
Статья
Google Scholar
Хейтманек М., Хейтманкова Н.: Телеоморфы и типы спаривания в комплексе Trichophyton mentagrophytes . Acta Univ Palacki Olomuc Fac Med. 1989, 123: 11-33.
PubMed
CAS
Google Scholar
Леннон М.Б., Сухадольник Р.Дж.: Биосинтез 3′-дезоксиаденозина с помощью Cordyceps militaris . Механизм редукции. Biochim Biophys Acta. 1976, 425: 532-536.
PubMed
CAS
Статья
Google Scholar
Reichard P: Рибонуклеотидредуктазы: субстратная специфичность аллостерией. Biochem Biophys Res Commun.2010, 396: 19-23. 10.1016 / j.bbrc.2010.02.108.
PubMed
CAS
Статья
Google Scholar
Лю М., Паначчоне Д.Г., Шардл К.Л.: Филогенетический анализ выявил монофилетическое происхождение гена алкалоида спорыньи dmaW в грибах. Evol Bioinform. 2009, 5: 15-30.
Google Scholar
Браун Д.В., Маккормик С.П., Александр Н.Дж., Проктор Р.Х., Дежарден А.Е.: Генетический и биохимический подход к изучению трихотеценового разнообразия у Fusarium sporotrichioides и Fusarium graminearum .Fungal Genet Biol. 2001, 32: 121-133. 10.1006 / fgbi.2001.1256.
PubMed
CAS
Статья
Google Scholar
Pöggeler S, Nowrousian M, Ringelberg C, Loros JJ, Dunlap JC, Kuck U. Анализ микроматрицы и ПЦР в реальном времени выявляет экспрессию генов, зависимую от типа спаривания, в гомоталлическом грибе. Mol Genet Genomics. 2006, 275: 492-503. 10.1007 / s00438-006-0107-у.
PubMed
Статья
Google Scholar
Kamata N, Sato H, Shimazu M: сезонные изменения в заражении куколок гусеницы бука, Quadriccarifera punctatella (Motsch.) (Lep., Notodontidae), по Cordyceps militaris Link (Clavicipitales, Clavicipitaceles) почва японского букового леса. J Appl Entomol. 1997, 121: 17-21. 10.1111 / j.1439-0418.1997.tb01364.x.
Артикул
Google Scholar
Clarkson JP, Staveley J, Phelps K, Young CS, Whipps JM: высвобождение аскоспор и выживаемость в Sclerotinia sclerotiorum .Mycol Res. 2003, 107: 213-222. 10.1017 / S0953756203007159.
PubMed
Статья
Google Scholar
Ohta Y, Lee JB, Hayashi K, Fujita A, Park DK, Hayashi T: In vivo противогриппозная активность иммуномодулирующего кислого полисахарида, выделенного из Cordyceps militaris , выращенного на проросших соевых бобах. J. Agric Food Chem. 2007, 55: 10194-10199. 10.1021 / jf0721287.
PubMed
CAS
Статья
Google Scholar
Sung GH, Poinar GO, Spatafora JW: самые старые ископаемые свидетельства паразитизма животных грибами подтверждают меловое разнообразие симбиозов грибков и членистоногих. Mol Phylogenet Evol. 2008, 49: 495-502. 10.1016 / j.ympev.2008.08.028.
PubMed
Статья
Google Scholar
Wang CS, St Leger RJ: Metarhizium anisopliae гомолог перилипина MPL1 регулирует липидный обмен, аппрессорное тургорное давление и вирулентность.J Biol Chem. 2007, 282: 21110-21115. 10.1074 / jbc.M6095.
PubMed
CAS
Статья
Google Scholar
Wang CS, Duan ZB, St Leger RJ: Осмосенсор MOS1 из Metarhizium anisopliae необходим для адаптации к гемолимфе насекомого-хозяина. Эукариотическая клетка. 2008, 7: 302-309. 10.1128 / EC.00310-07.
PubMed
PubMed Central
Статья
Google Scholar
Wang CS, St Leger RJ: Коллагеновая защитная оболочка позволяет Metarhizium anisopliae избегать иммунных реакций насекомых. Proc Natl Acad Sci USA. 2006, 103: 6647-6652. 10.1073 / pnas.0601951103.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Wang CS, St Leger RJ: Адгезин MAD1 из Metarhizium anisopliae связывает адгезию с образованием бластоспор и вирулентностью для насекомых, а адгезин MAD2 позволяет прикрепляться к растениям.Эукариотическая клетка. 2007, 6: 808-816. 10.1128 / EC.00409-06.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Freitag M, Williams RL, Kothe GO, Selker EU: Гомолог цитозинметилтрансферазы необходим для индуцированной повтором точечной мутации в Neurospora crassa . Proc Natl Acad Sci USA. 2002, 99: 8802-8807. 10.1073 / pnas.132212899.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Montiel MD, Lee HA, Archer DB: Доказательства RIP (точечная мутация, индуцированная повтором) в последовательностях транспозаз Aspergillus oryzae . Fungal Genet Biol. 2006, 43: 439-445. 10.1016 / j.fgb.2006.01.011.
PubMed
CAS
Статья
Google Scholar
Long M, Betran E, Thornton K, Wang W: Происхождение новых генов: взгляды молодых и старых. Nat Rev Genet. 2003, 4: 865-875.
PubMed
CAS
Статья
Google Scholar
Экман Д., Элофссон А. Идентификация и количественная оценка последовательностей орфанных белков в грибах. J Mol Biol. 2010, 396: 396-405. 10.1016 / j.jmb.2009.11.053.
PubMed
CAS
Статья
Google Scholar
Ohm RA, de Jong JF, Lugones LG, Aerts A, Kothe E, Stajich JE, de Vries RP, Record E, Levasseur A, Baker SE, Bartholomew KA, Coutinho PM, Erdmann S, Fowler T.J., Gathman AC, Lombard V, Henrissat B, Knabe N, Kües U, Lilly WW, Lindquist E, Lucas S, Magnuson JK, Piumi F, Raudaskoski M, Salamov A, Schmutz J, Schwarze FW, vanKuyk PA, Horton JS, Григорьев И.В. и др.: Геномная последовательность модельного гриба Schizophyllum commune .Nat Biotechnol. 2010, 28: 957-963. 10.1038 / nbt.1643.
PubMed
CAS
Статья
Google Scholar
Corson TW, Crews CM: Молекулярное понимание и современное применение традиционных лекарств: победы и испытания. Клетка. 2007, 130: 769-774. 10.1016 / j.cell.2007.08.021.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Yan WJ, Li TH, Lin QY, Song B, Jiang ZD: Оценка безопасности Cordyceps guangdongensis .Food Chem Toxicol. 2010, 48: 3080-3084. 10.1016 / j.fct.2010.08.001.
PubMed
CAS
Статья
Google Scholar
Хейтман Дж .: Эволюция эукариотических микробных патогенов посредством скрытого полового размножения. Клеточный микроб-хозяин. 2010, 8: 86-99. 10.1016 / j.chom.2010.06.011.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Кини S: Механизм и контроль инициации мейотической рекомбинации.Curr Top Dev Biol. 2001, 52: 1-53.
PubMed
CAS
Статья
Google Scholar
Masson JY, West SC: Рекомбиназы Rad51 и Dmc1: неидентичные близнецовые отношения. Trends Biochem Sci. 2001, 26: 131-136. 10.1016 / S0968-0004 (00) 01742-4.
PubMed
CAS
Статья
Google Scholar
Фишер Р.П .: Секреты двойного агента: CDK7 в контроле клеточного цикла и транскрипции.J Cell Sci. 2005, 118: 5171-5180. 10.1242 / jcs.02718.
PubMed
CAS
Статья
Google Scholar
Schatz MC, Delcher AL, Salzberg SL: Сборка больших геномов с использованием секвенирования второго поколения. Genome Res. 2010, 20: 1165-1173. 10.1101 / gr.101360.109.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Тер-Оганесян В., Ломсадзе А., Чернов Ю.О., Бородовский М. Прогнозирование генов в новых геномах грибов с использованием алгоритма ab initio с обучением без учителя.Genome Res. 2008, 18: 1979–1990. 10.1101 / gr.081612.108.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Zhang Z, Carriero N, Zheng D, Karro J, Harrison PM, Gerstein M: PseudoPipe: автоматизированный конвейер идентификации псевдогенов. Биоинформатика. 2006, 22: 1437-1439. 10.1093 / биоинформатика / btl116.
PubMed
CAS
Статья
Google Scholar
Сервер SignalP 3.0. [http://www.cbs.dtu.dk/services/SignalP/]
Сервер RepeastMasker. [http://www.repeatmasker.org/]
Benson G: Поиск тандемных повторов: программа для анализа последовательностей ДНК. Nucleic Acids Res. 1999, 27: 573-580. 10.1093 / nar / 27.2.573.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Кохани О., Джентлс А.Дж., Ханкус Л., Юрка Дж .: Аннотации, представление и проверка повторяющихся элементов в Repbase: RepbaseSubmitter и Censor.BMC Bioinformatics. 2006, 7: 474-10.1186 / 1471-2105-7-474.
PubMed
PubMed Central
Статья
Google Scholar
Schmidt HA, Strimmer K, Vingron M, von Haeseler A: TREE-PUZZLE: филогенетический анализ максимального правдоподобия с использованием квартетов и параллельных вычислений. Биоинформатика. 2002, 18: 502-504. 10.1093 / биоинформатика / 18.3.502.
PubMed
CAS
Статья
Google Scholar
Тейлор Дж. У., Берби М: Расхождения в датах на грибном древе жизни: обзор и новый анализ. Mycologia. 2006, 98: 838-849. 10.3852 / mycologia.98.6.838.
PubMed
Статья
Google Scholar
Lucking R, Huhndorf S, Pfister DH, Plata ER, Lumbsch HT: Грибки эволюционировали правильно. Mycologia. 2009, 101: 810-822. 10.3852 / 09-016.
PubMed
Статья
Google Scholar
Анализ и классификация белковых последовательностей InterPro. [http://www.ebi.ac.uk/interpro/]
База данных Pfam. [http://pfam.sanger.ac.uk/]
Шомбург Д., Шомбург Иллинойс: базы данных ферментов. Методы Мол биол. 2010, 609: 113-128. 10.1007 / 978-1-60327-241-4_7.
PubMed
CAS
Статья
Google Scholar
Сервер Cytochrome P450. [http://drnelson.uthsc.edu/CytochromeP450.html]
Saier MH, Tran CV, Barabote RD: TCDB: База данных классификации транспортеров для анализа и информации о мембранных транспортных белках. Nucleic Acids Res. 2006, 34: D181-186. 10.1093 / нар / gkj001.
PubMed
CAS
PubMed Central
Статья
Google Scholar
База данных GPCR. [http://www.gpcr.org/7tm/]
База PHI: Взаимодействие патоген-хозяин. [http: // www.phi-base.org]
Канехиса М., Гото С., Кавасима С., Накая А.: Базы данных KEGG в GenomeNet. Nucleic Acids Res. 2002, 30: 42-46. 10.1093 / nar / 30.1.42.
PubMed
CAS
PubMed Central
Статья
Google Scholar
SMURF. [http://www.jcvi.org/smurf/index.php]
Ананд С., Прасад М.В., Ядав Г., Кумар Н., Шехара Дж., Ансари М.З., Моханти Д.: SBSPKS: анализ последовательности на основе структуры поликетидсинтаз.Nucleic Acids Res. 2010, 38: W487-496. 10.1093 / нар / gkq340.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Kim YT, Lee YR, Jin J, Han KH, Kim H, Kim JC, Lee T, Yun SH, Lee YW: Для синтеза зеараленона в Gibberella zeae требуются два разных гена поликетидсинтазы. Mol Microbiol. 2005, 58: 1102-1113. 10.1111 / j.1365-2958.2005.04884.x.
PubMed
CAS
Статья
Google Scholar
Dao HP, Mathieu F, Lebrihi A: Две пары праймеров для обнаружения продуцентов OTA методом ПЦР. Int J Food Microbiol. 2005, 104: 61-67. 10.1016 / j.ijfoodmicro.2005.02.004.
PubMed
CAS
Статья
Google Scholar
Artigot MP, Loiseau N, Laffitte J, Mas-Reguieg L, Tadrist S, Oswald IP, Puel O: молекулярное клонирование и функциональная характеристика двух цитохромов P450 CYP619, участвующих в биосинтезе патулина в Aspergillus clavatus.Микробиология. 2009, 155: 1738-1747. 10.1099 / mic.0.024836-0.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Proctor RH, Desjardins AE, Plattner RD, Hohn TM: Ген поликетидсинтазы, необходимый для биосинтеза микотоксинов фумонизина в спаривающейся популяции Gibberella fujikuroi A. Fungal Genet Biol. 1999, 27: 100-112. 10.1006 / fgbi.1999.1141.
PubMed
CAS
Статья
Google Scholar
Shimizu T, Kinoshita H, Ishihara S, Sakai K, Nagai S, Nihira T: ген поликетидсинтазы, ответственный за биосинтез цитринина в Monascus purpureus . Appl Environ Microbiol. 2005, 71: 3453-3457. 10.1128 / AEM.71.7.3453-3457.2005.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Ehrlich KC, Chang PK, Yu J, Cotty PJ: Кластерный ген cypA биосинтеза афлатоксина необходим для образования G афлатоксина.Appl Environ Microbiol. 2004, 70: 6518-6524. 10.1128 / AEM.70.11.6518-6524.2004.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Yu JH, Leonard TJ: Для биосинтеза стеригматоцистина в Aspergillus nidulans требуется новая поликетид-синтаза типа I. J Bacteriol. 1995, 177: 4792-4800.
PubMed
CAS
PubMed Central
Google Scholar
Гардинер Д.М., Хоулетт Б.Дж.: Биоинформатический анализ и анализ экспрессии предполагаемого кластера биосинтетических генов глиотоксина Aspergillus fumigatus . FEMS Microbiol Lett. 2005, 248: 241-248. 10.1016 / j.femsle.2005.05.046.
PubMed
CAS
Статья
Google Scholar
Haese A, Schubert M, Herrmann M, Zocher R: Молекулярная характеристика гена энниатин синтетазы, кодирующего многофункциональный фермент, катализирующий образование N-метилдепсипептида в Fusarium scirpi .Mol Microbiol. 1993, 7: 905-914. 10.1111 / j.1365-2958.1993.tb01181.x.
PubMed
CAS
Статья
Google Scholar
Scott-Craig JS, Panaccione DG, Pocard JA, Walton JD: Циклическая пептид-синтетаза, катализирующая выработку HC-токсина в мицелиальных грибах Cochliobolus carbonum , кодируется открытой рамкой считывания длиной 15,7 килобаз. J Biol Chem. 1992, 267: 26044-26049.
PubMed
CAS
Google Scholar
Weber G, Schörgendorfer K, Schneider-Scherzer E, Leitner E: пептид-синтетаза, катализирующая выработку циклоспорина в Tolypocladium niveum , кодируется гигантской открытой рамкой считывания размером 45,8 килобаз. Curr Genet. 1994, 26: 120-125. 10.1007 / BF00313798.
PubMed
CAS
Статья
Google Scholar
Бреннер С., Джонсон М., Бриджем Дж., Голда Дж., Ллойд Д.Х., Джонсон Д., Ло С., МакКарди С., Фой М., Эван М., Рот Р., Джордж Д., Элетр С., Альбрехт Дж., Вермаас Э. , Williams SR, Moon K, Burcham T, Pallas M, DuBridge RB, Kirchner J, Fearon K, Mao J, Corcoran K: Анализ экспрессии генов путем массового параллельного секвенирования сигнатур (MPSS) на массивах микрогранул.Nat Biotechnol. 2000, 18: 630-634. 10.1038 / 76469.
PubMed
CAS
Статья
Google Scholar
Бруно В.М., Ван З., Марджани С.Л., Ойскирхен Г.М., Мартин Дж., Шерлок Дж., Снайдер М.: Исчерпывающая аннотация транскриптома грибкового патогена человека Candida albicans с использованием RNA-seq. Genome Res. 2010, 20: 1451-1458. 10.1101 / gr.109553.110.
PubMed
CAS
PubMed Central
Статья
Google Scholar
Benjamini Y, Yekutieli D: Контроль ложного обнаружения при множественном тестировании в зависимости. Ann Stat. 2001, 29: 1165-1188.